
Вирус который вызывает лимфому
Обновлено: 23.04.2024
Инфекционные причины лейкозов - лимфома Беркитта, иммунобластная лимфома, Т-клеточный лейкоз взрослых (ТКЛВ)
В экспериментальной лейкозологии существует много видов вирусов, с помощью которых индуцируют различные формы лейкозов. Известны также вирусы, вызывающие лейкозы у разных видов животных в естественных условиях. Для этих вирусов доказаны вертикальная трансмиссия (передача от матери потомству) и значительное распространение носительства в популяциях животных без признаков заболевания.
Эндемические формы инфекционных гемобластозов
Для большинства эндемических форм гемобластозов человека с определенностью установлено участие в индукции заболевания инфекционных агентов. В отношении некоторых убиквитарных форм существуют лишь более или менее убедительные подозрения. Рабочая группа МАИР в настоящее время причисляет к доказанным этиологическим факторам (группа 1) лимфопролиферативных гемобластозов следующие инфекционные агенты: вирус Эпштейна—Барр (EBV), вирус Т-клеточной лимфомы/лейкоза взрослых (HTLV-1), вирус иммунодефицита человека ВИЧ (HIV) и бактерию Helicobacter pylori.
Носительство EBV распространено очень широко и охватывает до 90 % взрослого населения. Вирус крайне неустойчив во внешней среде, поэтому заражение возможно только при прямом контакте — воздушно-капельным путем через слюну. Возраст, в котором происходит инфицирование, зависит от социальных условий: в развитых странах до трехлетнего возраста инфицируется около 20 % детей, тогда как в развивающихся —до 70 %. EBV обладает высокой тропностью к эпителию ротоглотки, где в нормальных условиях происходит его репродукция, и к В-лимфоцитам, в которых осуществляется бессимптомное носительство в виде кольцевой ДНК (эписома).
Доля содержащих эписому В-лимфоцитов невелика и довольно постоянна — 1 клетка на 10 5 —10 6 . В продуктивной фазе вирусная ДНК в В-клетках приобретает линейную форму.
В экваториальной зоне Африки с EBV связана лимфома Беркитта (ЛБ), которая занимает первое место по распространенности среди гемобластозов у детей на территориях, расположенных до 1550 м над уровнем моря в теплом и влажном климате. Особенно часто заболевание развивается у детей 5—13 лет; в 95 % всех случаев болезнь возникает до 16 лет. У детей моложе 2 лет заболевание практически не встречается. Классическое начало ЛБ: быстро растущая опухоль челюсти или органов брюшной полости у ребенка 5—8 лет. Мальчики преобладают в соотношении (1,7—2,0):1. Неберкиттовские лимфомы в этом регионе исключительно редки.
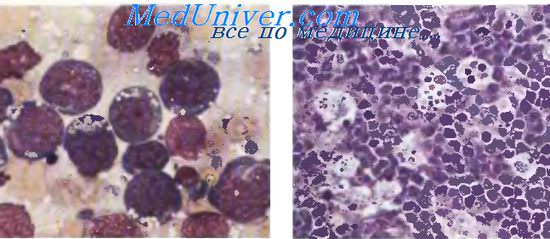
Морфология лимфомы Беркитта
Повышенное внимание к признакам лимфомы Беркитта (ЛБ) привело к тому, что это новообразование стали обнаруживать в западных странах среди НХЛ детского возраста (до 20 %). Раболеваемость в Африке значительно выше — в среднем 8 на 100 тыс. детей против 0,2 среди белого населения США.
Практически у 100 % больных лимфомой Беркитта (ЛБ) в африканских очагах в клетках опухоли выявляются включения ДНК EBV и определяются высокие титры антител к капсидным и ранним антигенам. Для здоровых детей с антителопродукцией к EBV риск ЛБ повышен в 50—60 раз. Среди спорадических случаев ЛБ носительство ДНК EBV в клетках опухоли много реже — около 30 %. В отличие от общего пула В-клеток ДНК ЕВУ содержится во всех опухолевых клетках ЛБ, причем в моноклоновой форме, что является основным подтверждением этиологической роли вируса. Известна способность EBV к трансформации клетки и индукции бесконечного деления В-лимфоцитов in vitro, тем не менее его роль в патогенезе ЛБ не вполне ясна.
Есть данные, что кофактором в развитии заболевания служит малярийный плазмодий: ареал высокого риска ЛБ совпадает, как правило, с эндемическими регионами малярии. У переселившихся в эти регионы из благополучных районов повышается вероятность возникновения обоих заболеваний. Проведение тотальной лекарственной профилактики малярии приводило к достоверному снижению заболеваемости ЛБ, а после прекращения противомалярийных мероприятий заболеваемость возвращалась на прежний уровень. Эпидемиологические наблюдения позволили построить следующую схему патогенеза ЛБ: малярийная инвазия, хроническая инфекция и, возможно, другие факторы могут служить активаторами пролиферации лимфоцитов.
Контакт с EBV вводит В-лимфоцит в бесконечный цикл деления (нечто близкое к инфекционному мононуклеозу). На следующем этапе случайная хромосомная транслокация способна вывести одну из активированных клеток из-под контроля, и она становится родоначальницей клона.
Анализ 665 случаев лимфомы Беркитта (ЛБ) из регистра Ибадана (Нигерия) показал значимое снижение относительной частоты заболеваний в период 1973—1990 гг. (37,1 % от всех опухолей у детей) по сравнению с 1960—1972 гг. (51,5 %). По всей видимости, это отражает реальное снижение заболеваемости за счет улучшения условий жизни и успешного контроля малярии.
Т-клеточный лейкоз взрослых (ТКЛВ) — заболевание, характерное для юго-восточных областей Японии (Окинава, Киуши, Шикоку), Экваториальной Африки и Карибских островов, Южной Америки и Ближнего Востока. В эндемических очагах до 20 % населения продуцируют антитела к HTLV-1 (среди населения США — 0,025 %). Наиболее распространен HTLV-1 в Африке —до 10 млн инфицированных, всего в мире их 15 — 20 млн. Трансмиссия вируса происходит вертикально (при грудном вскармливании ребенка), а также при гемотрансфузиях. Выявляемость носителей увеличивается с возрастом и достигает пика к 50 годам, причем доля женщин больше. Один случай ТКЛВ в год возникает среди приблизительно 1500 носителей HTLV-1.
В итоге заболевание развивается у 1—4 % носителей через 20— 30 лет после заражения, причем более 90 % заболевших серопозитивны к HTLV-1. Максимальное число заболевших регистрируется в Японии в возрасте около 60 лет, а в Африке — в 40—45 лет. Случаи заболевания детей исключительно редки, частота заболеваний у мужчин и женщин примерно равная. Смертность от ТКЛВ среди инфицированных достигает у мужчин 68, а у женщин 36 на 100 тыс.; среди серонегативных лиц она много ниже. Обследованные в эндемических районах матери заболевших ТКЛВ оказались инфицированными HTLV-1 в 100 %, тогда как матери больных другими формами лимфом — в 30 % случаев.
В Африке выявлено широкое распространение родственного вируса HTLV-2, который подозревается в инициации В-клеточных лейкозов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Информацию из данного раздела нельзя использовать для самодиагностики и самолечения. В случае боли или иного обострения заболевания диагностические исследования должен назначать только лечащий врач. Для постановки диагноза и правильного назначения лечения следует обращаться к Вашему лечащему врачу.
Лимфомы: причины появления, симптомы, диагностика и способы лечения.
Определение
Лимфомы представляют собой группу злокачественных опухолевых заболеваний лимфатической ткани, которая в нормальных условиях отвечает за иммунитет. Одним из первых клинических симптомов лимфом является увеличение лимфатических узлов разных локализаций.
Лимфомы, как и другие злокачественные опухоли, обладают склонностью к метастазированию, то есть миграции опухолевых клеток через кровь и лимфу, а также контактным путем в интактные (здоровые) ткани с последующим размножением в них.
Практически все виды лимфогранулематоза, или лимфомы Ходжкина, и около 65% неходжскинских лимфом проявляются безболезненным увеличением лимфатических узлов - 2 см и более. Однако диагноз того или иного вида лимфомы устанавливают исключительно на основании морфологического исследования биопсийного материала, взятого из опухоли.
Заболеваемость лимфомой Ходжкина в России составляет около 2 случаев на 100 тыс. населения в год, а неходжкинскими лимфомами – 5–7 случаев на 100 тыс.
Причины появления лимфом
Существует несколько возможных причин развития лимфом. Важно понимать, что в основе развития любого злокачественного заболевания, в том числе и лимфом, лежит нарушение функций клеток.
Способность к неконтролируемому росту опухоли возникает при нарушении естественного процесса деления клетки: в норме клетка делится ограниченное количество раз, созревает, выполняет определенную функцию, после чего заканчивает свою жизнь.
Некоторые вирусы могут напрямую влиять на ДНК лимфоцитов и способствовать их превращению в злокачественные клетки.
Лимфоциты делятся на два подвида: Т- и В-клетки (Т- и В-лимфоциты). Заражение Т-клеточным лимфотропным вирусом человека (HTLV-1) повышает риск возникновения у пациента определенных типов Т-клеточной лимфомы. Вирус передается половым путем и через кровь, а также от матери к ребенку через грудное молоко.
Вирусы, ослабляющие иммунологический надзор, косвенно являются причиной развития некоторых типов лимфом.
Некоторые инфекции могут стать причиной развития лимфомы из-за постоянной активации иммунной системы. При хронических инфекциях вырабатывается огромное количество лимфоцитов, что повышает вероятность возникновения мутаций во время деления клеток и ведет к риску формирования лимфомы. К подобным инфекциям относят Helicobacter pylori (часто встречается при хроническом гастрите и язвенной болезни желудка и двенадцатиперстной кишки), Chlamydophila psittaci (риск развития лимфомы в тканях глаза), вирусы гепатита В и С при длительном инфицировании.
К неинфекционным факторам риска развития лимфом относят:
- Воздействие химических веществ. Многочисленные исследования доказали, что влияние гербицидов, инсектицидов, бензола связано с повышенным риском развития лимфом.
- Наличие аутоиммунных заболеваний (ревматоидного артрита, системной красной волчанки и т.д.) является значимым фактором риска развития лимфом.
- Радиационное облучение оказывает большое влияние на скорость возникновения мутаций клеток и их озлокачествление.
- Генетические факторы. Наличие близкого родственника, страдающего лимфомой, значительно увеличивает риск.
- Врожденная недостаточность иммунитета при некоторых заболеваниях.
- Возраст. Некоторые типы лимфом развиваются у детей и подростков, а другие, напротив, у лиц старше 60 лет.
- Грудные импланты в редких случаях способствуют развитию одного из типов крупноклеточной лимфомы молочных желез.
Для лимфомы Ходжкина характерно наличие специфических клеток Рид–Березовского–Штернберга в пораженных лимфатических узлах, селезенке, печени или костном мозге.
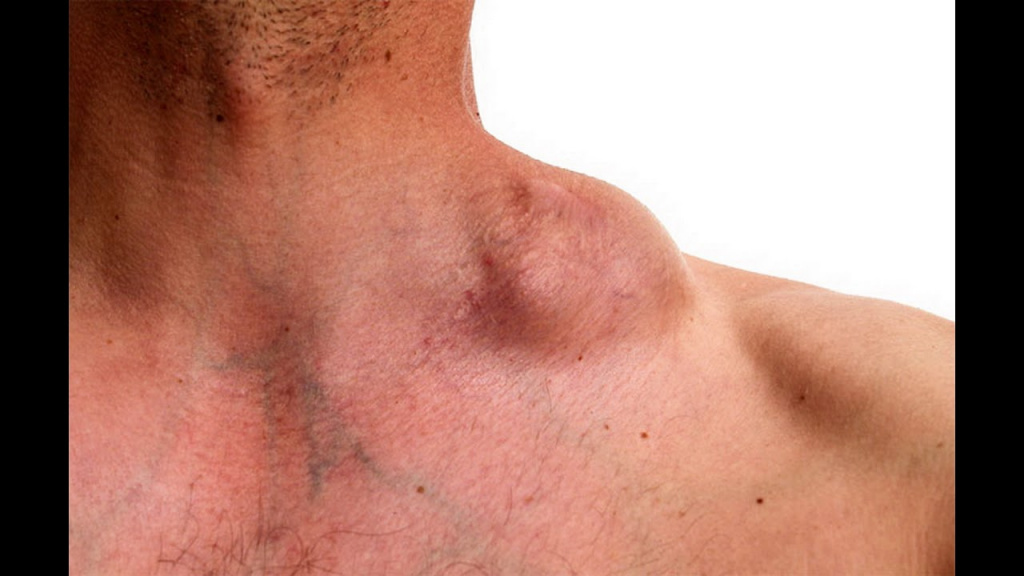
Увеличенные шейные лимфоузлы у пациента с лимфомой Ходжкина
К группе неходжкинских лимфом относятся:
- фолликулярная лимфома;
- лимфома маргинальной зоны;
- диффузная крупноклеточная В-клеточная лимфома;
- лимфома из клеток мантии;
- лимфома Беркитта;
- первичная лимфома центральной нервной системы;
- нодальные Т-клеточные лимфомы;
- первичные кожные лимфомы;
- неходжкинские лимфомы у больных, инфицированных ВИЧ;
- неходжкинские лимфомы у больных, инфицированных вирусами гепатита В и С.
Самый ранний симптом – увеличение лимфатических узлов на фоне полного благополучия. Увеличенные лимфоузлы безболезненны.
Различные клинические проявления зависят от локализации опухоли: если происходит рост опухоли в лимфатических узлах грудной клетки, возможно сдавливание дыхательных путей и появление охриплости голоса, нарушение глотания, затруднение дыхания, навязчивый кашель.
При расположении лимфомы в средостении появляется синдром сдавления верхней полой вены: возникают головные боли, отек и расширение вен лица и шеи.
В некоторых случаях присутствуют только общие симптомы: потеря аппетита, снижение массы тела, повышение температуры тела, ночная потливость, слабость, иногда – кожный зуд и боль в увеличенных лимфоузлах.
На фоне снижения иммунитета могут присоединяться инфекционные заболевания - часто активируется вирус ветряной оспы, проявляющийся опоясывающим герпесом.
Описаны случаи поражения органов брюшной полости (проявляется увеличением объема живота за счет роста опухоли и скопления жидкости, появлением болей, желтухи, тошноты и рвоты) и костного мозга (боли в костях, спине, частые переломы, бледность, повышенная кровоточивость).
Диагностика лимфом
Диагноз того или иного вида лимфомы устанавливают исключительно на основании морфологического исследования биопсийного материала (кусочка или целого лимфоузла, полученного в ходе небольшой операции), взятого из опухоли (гистологические, цитогенетические и молекулярно-генетические методы исследования). Дополнительные исследования врач назначит для диагностики стадии заболевания и его осложнений. Список исследований зависит от предположительной локализации опухоли и может быть изменен.
-
Клинический анализ крови.
Синонимы: Общий анализ крови, ОАК. Full blood count, FBC, Complete blood count (CBC) with differential white blood cell count (CBC with diff), Hemogram. Краткое описание исследования Клинический анализ крови: общий анализ, лейкоформула, СОЭ См. также: Общий анализ – см. тест № 5, Лейкоцит.

Обзор
Автор
Редактор
Обратите внимание!
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science.
Исследования в области происхождения злокачественных опухолей, ежегодно уносящих миллионы человеческих жизней, ведутся с XIX века. Работая в области молекулярной вирусологии, Вармус совместно с Дж. М. Бишопом в исследованиях 1970-х гг. сделали открытие, которое по-новому осветило долго остававшуюся дискуссионной проблему этиологии опухолей у человека и животных. Согласно полученным результатам, неконтролируемый рост клеток, образующих опухоль, вызывается не только проникающим в клетку извне онковирусом, но и внутренними процессами в самой клетке. Вармус доказал, что нормальные гены роста клетки вследствие случайных спонтанных мутаций под воздействием химических канцерогенов или, порой, процесса старения могут изменять свою молекулярную структуру и таким образом превращаться в протовирусы онкогенной природы. За открытие клеточного происхождения онкогенных протовирусов Вармус совместно с Дж. М. Бишопом в 1989 г. были удостоены Нобелевской премии по физиологии и медицине [1].
Один из самых распространённых вирусов в человеческой популяции — вирус Эпштейна-Барр (рис. 1) — был открыт и описан в 1964 году двумя английскими вирусологами: Майклом Эпштейном и Ивонной Барр. Вирус Эпштейна-Барр (ВЭБ) — член семейства герпесвирусов [6, 7]. В инфицированных клетках вирусная ДНК, как правило, не встроена в клеточный геном, а находится в ядре в виде замкнутого кольца (эписомы). Биологическое значение интеграции ВЭБ в геном клетки остается неясным. Высказываются предположения, что эписомная ДНК необходима для реализации полноценной репликации ВЭБ, завершающейся формированием вирусных частиц [8].

В отличие от многих других герпесвирусов, вирус Эпштейна-Барр поражает в первую очередь эпителиальные клетки ротовой полости, глотки, миндалин. Здесь он размножается наиболее активно, и поэтому главным путём заражения вирусом являются поцелуи (вот и приехали). Наибольшее количество вирусных частиц находится в клетках эпителия около слюнных желез, и со слюной выделяется большое количество их. Не удивительно, что инфекционный мононуклеоз — самое распространённое заболевание, вызываемое вирусом Эпштейна-Барр, — называют ещё болезнью поцелуев [9].
После первой встречи человека с ВЭБ вирус в незначительном количестве сохраняется в организме хозяина в течение всей жизни. Однако если любой элемент иммунного ответа нарушен, даже незначительное количество ВЭБ-инфицированных клеток может преумножиться колоссально [12].
Инфицированные В-клетки могут значительное время находиться в миндалинах, что позволяет вирусу выделяться во внешнюю среду со слюной. С зараженными клетками ВЭБ распространяется по другим органам. В пораженных вирусом клетках возможно два вида развития: литический, приводящий к разрушению клетки-хозяина, и латентный (клетка заражена, но ничего не выдает нахождения вируса), когда число вирусных копий небольшое и клетка не разрушается. ВЭБ может длительно находиться в В-лимфоцитах, эпителии носоглотки и слюнных железах. Кроме того, он способен проникать и в другие клетки: Т-лимфоциты, NK-клетки, макрофаги, нейтрофилы, эпителиоциты сосудов (рис. 2а, 2б) [13].

Злокачественные лимфомы (например, увеличение лимфоузлов), согласно данным Международного агентства по изучению рака, составляют 3–4% среди всех регистрируемых в мире злокачественных новообразований [14]. Лимфомы делят на две основные группы: лимфома Ходжкина (20–30 % всех лимфом) и неходжкинские лимфомы (около 70%) [14, 15].
Неходжикинские лимфомы — это совокупность новообразований, в возникновении которых принимают участие различные агенты [18]. Первая группа — это вирусы, трансформирующие лимфоциты и другие клетки (ВЭБ, HHV-8). Вторая группа представлена факторами различной природы, вызывающими иммунодефицитные состояния. К таким факторам в первую очередь относится ВИЧ (вирус иммунодефицита человека), вызывающий у инфицированного лица подавление иммунитета в результате истощения пула Т-лимфоцитов CD4+ и возникновение СПИДа. В третью группу входят некоторые инфекции, (например, H. pylori), которые увеличивают риск возникновения лимфом на фоне вызываемой ими хронической стимуляции иммунной системы и постоянной активации лимфоцитов [17].
Исследования последних лет свидетельствуют о том, что в патогенезе ВЭБ-ассоциированных патологий чрезвычайно важную роль играет вредный LMP1 — латентный мембранный белок 1, кодируемый одноименным геном (LMP1). Он обладает свойствами онкобелка и функционирует как постоянно активный псевдорецептор. Он способен изменять В-лимфоциты человека [19].
Есть предположение, что аминокислотные замены, накапливающиеся в LMP1, по-видимому, также вносят свой вклад в возникновение опухолей. Механизм этого процесса окончательно не установлен, но предполагается, что усиленный трансформирующий эффект мутированных LMP1 может представлять важную составляющую этого процесса. При этом показано, что различия в последовательности гена LMP1 могут определять агрессивный географически локализованный генотип ВЭБ [21].
Из известных механизмов действия LMP-2 (второй из братьев семейства LMP), расположенного на противоположном конце линейного генома, упоминается лишь способность этих белков совместно повышать сигнальную трансдукцию в ВЭБ (+) клетках [19].
По данным Харальда цур Хаузена, связь между вирусом и раковым заболеванием считается установленной при определении следующих критериев:
- эпидемиологические доказательства того, что вирусная инфекция является фактором риска для развития специфической опухоли;
- присутствие и сохранение генома вируса в клетках опухоли;
- стимуляция пролиферации клеток после введения генома (или его части) вируса в ткани культуры клеток;
- демонстрация того, что геном возбудителя индуцирует пролиферацию и злокачественный фенотип опухоли [19].
Однако канцерогенность ВЭБ далеко не однозначна. Несмотря на то, что кодируемые вирусом продукты способны вызывать пролиферацию инфицированных клеток, ведущую к возникновению лимфом у больных с иммунодефицитом, эти клинически агрессивные опухоли довольно часто поликлональны и подвергаются регрессии при восстановлении иммунного ответа на ВЭБ. Такие опухоли как лимфома Беркитта (ЛБ) и лимфома Ходжкина (ЛХ) встречаются не только в ВЭБ-ассоциированных, но и в ВЭБ-неассоциированных вариантах, что говорит о том, что патогенез этих новообразований связан не только с ВЭБ. Кроме того, злокачественные клетки больных ЛБ и ЛХ отличаются фенотипически от клеток ЛКЛ, полученных под воздействием ВЭБ in vitro, и не экспрессируют ряд белков, необходимых для трансформирующего роста. Эти находки позволяют предположить, что опухолевые клетки могут возникать и под воздействием факторов невирусного происхождения, а также зависеть от различных усиливающих рост клеток стимулов [17].
Лабораторная диагностика ВЭБ-инфекции базируется на цитологическом исследовании крови или костного мозга, серологических исследованиях и ПЦР. С помощью метода ПЦР можно определить ДНК вируса в плазме до клинических проявлений болезни, а репликация вируса в организме является показанием к противовирусной терапии и критерием эффективности проведенного лечения. Материалом для исследования служат слюна или рото- и носоглоточная слизь, соскоб эпителиальных клеток урогенитального тракта, кровь, спинномозговая жидкость, ткани опухоли и костный мозг. Как у больных ВЭБ, так и у носителей может быть получен положительный результат в ПЦР. Поэтому для их дифференцировки проводится количественный ПЦР-анализ для определения количества копий вирусного генома. У маленьких детей (до 1–3-х лет) по причине недостаточно сформированного иммунитета диагностика по антителам затруднительна, поэтому в данной группе пациентов в помощь приходит именно ПЦР. Однако в силу того, что ПЦР-анализ информативен только при размножении (репликации) вируса, то существует и определенный процент ложноотрицательных результатов (до 30%), связанный именно с отсутствием репликации в момент исследования. При этом важно сопоставление результатов клинических, серологических и молекулярных обследований в определении ВЭБ-инфекции, как причины имеющегося заболевания [1].
Специфическая профилактика (вакцинация) против ВЭБ не разработана, но проводятся клинические испытания. Основной проблемой при разработке вакцины является большое отличие в белковом составе вируса на разных фазах его существования. Впрочем, в настоящее время разрабатывается вакцина, которая содержит рекомбинантный поверхностный антиген gp350. После вакцинации первичная инфекция протекает субклинически, но собственно инфицирование человека не предупреждается. Кроме того, вырабатывающиеся нейтрализующие антитела не влияют на течение различных форм латентной инфекции, в том числе опухолей. Профилактические меры сводятся к укреплению иммунитета, закаливанию детей, мерам предосторожности при появлении больного в окружении, соблюдение правил личной гигиены.
Заключение
Широкое распространение ВЭБ с выраженным трансформирующим потенциалом среди населения планеты и редкого возникновения в инфицированной популяции связанных с этим вирусом опухолей с преимущественной их локализацией в определенных географических регионах позволяет сделать важный вывод. Подобно большинству опухолей иной вирусной природы, в патогенезе ВЭБ-ассоциированных новообразований важную роль играют дополнительные факторы, и одного ВЭБ недостаточно для возникновения опухоли. ВЭБ лишь инициирует пролиферацию инфицированных им клеток, а последующие события влияют на гистопатологический спектр возникающих неоплазий. Одним из важнейших факторов, в значительной степени определяющих возникновение ВЭБ-ассоциированных опухолей, служит выраженная иммуносупрессия (врожденная, ятрогенная или индуцированная любой вирусной инфекцией, и в первую очередь ВИЧ), приводящая к утрате функции иммунного распознавания клеток, инфицированных ВЭБ.
Таким образом, несмотря на многолетнее изучение связи ВЭБ с опухолями человека, вопрос о роли вируса в их возникновении до конца не изучен. Раскрытие механизма злокачественной трансформации вирусом, персистирующим в латентном состоянии более чем у 90% населения планеты, — задача чрезвычайно сложная. Однако технические достижения последних лет, существенно повысившие специфичность исследований, позволяют надеяться, что детали ВЭБ-ассоциированного канцерогенеза будут выяснены.

Обзор
Светооптическая микроскопия ткани, пораженной диффузной В-клеточной крупноклеточной лимфомой.
Автор
Редакторы

Партнер номинации — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Генеральный партнер конкурса — международная инновационная биотехнологическая компания BIOCAD.
Введение
Что ж, теперь я хочу пригласить вас в удивительный и сложный мир молекулярного и генетического разнообразия лимфомы. Уверяю вас, что биология рака — как огромный океан: себя можно представить в небольшом батискафе в бездонном морском пространстве; здесь много неизвестного, ты порой не понимаешь, как оценить весь масштаб происходящего, ощущаешь себя мелкой крупицей. Все это пугает и завораживает одновременно. Готовы к погружению?
Такая разная лимфома
Два паттерна экспрессии генов ДВККЛ определяют клинический прогноз
Рисунок 1. Кривые общей выживаемости больных диффузной В-крупноклеточной лимфомой в зависимости от молекулярного подтипа.
Возможно, именно различия в экспрессии генов лежат в основе различного исхода пациентов? Авторы этого же исследования проанализировали общую выживаемость больных, которые получали сопоставимые схемы лечения. Оказалось, что пациенты с GCB-подтипом ДВККЛ имели статистически значимые различия в выживаемости в сравнении с АВС-подтипом (activated B-cell like subtype) ДВККЛ: в среднем общая пятилетняя выживаемость для всех пациентов составила 52%, однако 76% пациентов с GCB-подтипом по достижении пятилетнего рубежа были все еще живы, в то время как только 16% пациентов с ABC-подтипом остались в живых [3]. Исследовательская группа предоставила данные по выживаемости в виде кривых выживаемости Каплана—Мейера (рис. 1). Эти кривые имеют ступенчатый вид и очень часто используются в клинических исследованиях для оценки выживаемости пациентов. В таких графиках каждая ступенька означает наступление неблагоприятного для пациента события: прогрессирование заболевания (при изучении выживаемости без прогрессирования) или летальный исход (при изучении общей выживаемости).
Таким образом, был получен окончательный вывод: диффузная В-клеточная крупноклеточная лимфома представляет собой не одно заболевание, а группу различных по молекулярному патогенезу заболеваний с неодинаковым клиническим прогнозом.
К большому сожалению, даже на сегодняшний день в рутинной онкогематологической практике использование методов анализа экспрессии генов невозможно из-за дороговизны и сложности метода. В качестве альтернативного метода определения молекулярного подтипа ДВККЛ в клинике врачами-патологоанатомами используется окрашивание ткани опухоли в исследовании, основанном на взаимодействии антитела с антигеном — иммуногистохимическая реакция по алгоритму Hans [4], [5]. Данный метод основан на оценке относительного уровня экспрессии всего трех маркеров: CD10, BCL6 и MUM1 (рис. 2). Хоть этот метод более доступен, он не так точен и не позволяет в чистом виде выделить АВС-подтип, поэтому в иммуногистохимическом алгоритме предусмотрено разделение ДВККЛ на GCB- и non-GCB-подтипы.
Рисунок 2. Алгоритм Hans для иммуногистохимической идентификации подтипов.
Молекулярные основы патогенеза ABC- и GCB-подтипов ДВККЛ
Генетические поломки, обуславливающие формирование злокачественных лимфом, возникают на определенном этапе нормального развития В-лимфоцита. Опухолевые В-лимфоциты неходжкинских лимфом имеют биологические особенности нормальных В-лимфоцитов, и они используют эти свойства для собственного выживания и размножения злокачественного клона. Это означает, что функционирование В-клеток лимфом в значительной степени определяется уровнем развития или дифференцировки В-лимфоцитов, из которых опухоль происходит. Обратите внимание на основные последовательные стадии дифференцировки В-лимфоцита (рис. 3), начиная от стволовой кроветворной клетки и заканчивая плазматической клеткой, основная функция которой — секреция защитных антител [6]. Это знание еще пригодится нам в дальнейшем.
Рисунок 3. Последовательные этапы развития В-лимфоцита.
В костном мозге в процессе В-лимфопоэза (формирования В-лимфоцита из стволовой кроветворной клетки) происходит созревание В-клеточного рецептора лимфоцита — будущего специфичного антитела — за счет V(D)J-рекомбинации при участии ферментов RAG1 и RAG2, которые вносят разрывы в двухцепочечную молекулу ДНК, устраняемые простым соединением концов ДНК. Данное молекулярное событие может стать источником формирования хромосомных транслокаций (перестроек) и образования химерных генов, которые в нормальных клетках отсутствуют и гиперэкспрессия которых значима в патогенезе лимфом. Другим потенциальным источником возникновения генетических поломок является герминативный центр лимфатического фолликула, о котором мы уже говорили ранее. Именно здесь происходят события, связанные с модификацией ДНК, которые представляют собой еще один источник мутагенеза — соматическая гипермутация (накопление большего количества мутаций в генах иммуноглобулинов при участии цитозиндезаминазы AID), в процессе которой происходит созревание аффинитета (сродства) В-клеточного рецептора к антигену, и переключение классов иммуноглобулинов. Таким образом, на определенных этапах нормального развития любого В-лимфоцита существуют молекулярные механизмы, которые создают предпосылки к возникновению генетических поломок и, как следствие, — к развитию лимфом. Возникновение той или иной генетической поломки в В-лимфоците носит вероятностный характер.
Рисунок 4. В-клеточная дифференцировка, лимфомагенез и профиль экспрессии генов на различных этапах созревания В-лимфоцита. Злокачественные лимфомы могут возникнуть на разных этапах нормального развития В-лимфоцита. По мере созревания В-лимфоцита его генетическая регуляторная программа переключается с профиля В-лимфоцита герминативного центра на плазмоцитоидный профиль.
Наконец, мы добрались до кульминационной части данного раздела. В случае прогностически более благоприятного варианта — GCB-подтипа ДВККЛ — экспрессионный профиль генов таков, что он соответствует профилю В-лимфоцита герминативного центра, при этом активность молекулярного сигнального пути NF-kB (nuclear factor kappa-B) в таких клетках снижена [7]. Наоборот, в случае клинически неблагоприятного АВС-подтипа ДВККЛ активность NF-kB высока, что может служить одним из механизмов избегания программируемой клеточной гибели — апоптоза. Такие опухолевые клетки экспрессируют маркеры, характерные для плазматических клеток, включая IRF4 и транскрипционный фактор XBP1 — регулятор секреции иммуноглобулинов. Однако при этом полное развитие В-лимфоцитов АВС-подтипа лимфомы в нормальный плазмоцит невозможно из-за блока дифференцировки [7].
Роль активации NF-kB и сигнального пути В-клеточного рецептора в АВС-подтипе ДВККЛ
Как уже было отмечено ранее, успешная стратегия выживания опухолевого В-лимфоцита может объясняться высокой зависимостью от NF-kB для АВС-подтипа ДВККЛ, но не для GCB-подтипа. А также известно о том, что каскадный внутриклеточный сигнальный путь В-клеточного рецептора в случае АВС-подтипа находится в состоянии хронической гиперактивации [7]. Существует довольно несложный способ продемонстрировать, что эти сигнальные пути являются жизненно важными для опухолевых клеток. Для этого достаточно их заблокировать или выключить. На клеточных линиях, являющихся моделями АВС-подтипа ДВККЛ, исследователи Davis R.E. и соавторы (2010) с помощью явления РНК-интерференции продемонстрировали нежизнеспособность опухолевого клона при подавлении экспрессии одного из ключевых посредников сигнала от В-клеточного рецептора — Брутоновской тирозинкиназы [8]. С другой стороны, Lam L.T. и соавторы (2005) успешно подтвердили токсический эффект блокатора IKK, который связан с путем NF-kB в В-лимфоцитах АВС-подтипа [9].
Что, если результаты этих исследований использовать в терапевтических целях? Сложная система внутриклеточных молекул-посредников злокачественного В-лимфоцита АВС-подтипа ДВККЛ может стать интересным объектом терапевтического воздействия (рис. 5). В фармакологической тяжелой артиллерии врача-гематолога уже сегодня имеются блокаторы В-клеточного рецепторного каскада (ибрутиниб — ингибитор Брутоновской тирозинкиназы, дувелисиб — ингибитор PI3K, полатузумаб ведотин — связывает CD79b) и протеасомные ингибиторы (бортезомиб), которые, косвенно воздействуя на NF-kB, предотвращают его проникновение в клеточное ядро и реализацию программы выживания. Более интересной терапевтической мишенью для лечения ДВККЛ могут стать компоненты комплекса CARD11/BCL10/MALT1 (рис. 5). Приблизительно 10% всех АВС-подтипов лимфом имеют активационную мутацию в CARD11, которая обуславливает автономное формирование внутрицитоплазматических белковых агрегатов CARD11/BCL10/MALT1/TRAF6, что приводит к активации IKK, а в последующем — к запуску NF-kB [7]. Причем такая мутация делает независимым сигнальный путь NF-kB от посредников (мессенджеров) сигнала от В-клеточного рецептора, стоящих выше CARD11.
Рисунок 5. Функционирование внутриклеточного сигнального каскада В-клеточного рецептора для нормального В-лимфоцита (А) и в случае В-лимфоцита АВС-подтипа ДВККЛ с мутантным и диким вариантами CARD11 (B).
Повторяющиеся мутационные паттерны ДВККЛ отражают профиль экспрессии генов
Рисунок 6. Секторальные диаграммы, отражающие относительное представительство молекулярного подтипа ДВККЛ на основе профиля экспрессии генов одному из четырех генетических подтипов, и наоборот.
(а) — доля молекулярных подтипов ДВККЛ (АВC-подтип, GCB-подтип и неклассифицируемый) среди генетических вариантов ДВККЛ (MCD, BN2, N1, EZB).
(б) — доля генетических подтипов ДВККЛ (MCD, BN2, N1, EZB) среди молекулярных подтипов ДВККЛ (АВC-подтип, GCB-подтип и неклассифицируемый). (в) — общая доля всех генетических подтипов ДВККЛ среди всех случаев.
Не так давно представления о разнообразии ДВККЛ несколько усложнились. Schmitz et al. (2018) провели секвенирование экзома [10] и транскриптомный анализ RNA-seq 574 образцов ДВККЛ и выявили группы повторяющихся от случая к случаю мутаций в опухолевой ДНК, которые ассоциированы с одним из молекулярных подтипов [11]. Исследовательской группе удалось выделить четыре генетических типа ДВККЛ: MCD (сосуществующие мутации MYD88 и CD79b в опухоли), BN2 (транслокация BCL6 и мутация NOTCH2), N1 (мутация NOTCH1) и EZB (мутация EZH2 и транслокация BCL2) [11]. Только 46,6% случаев ДВККЛ удалось классифицировать по данным генетическим группам; оставшаяся часть лимфом не может быть отнесена ни к одному из подтипов и считается неклассифицированной. Нетрудно заметить (рис. 6), что подавляющее большинство молекулярных подтипов в случае MCD и N1 генетических вариантов соответствуют АВС-подтипу, а EZB вариант — GCB-подтипу [11]. Примечательно, что определение генетических групп, подобно молекулярным группам, имеет клиническое значение с точки зрения прогноза для пациента. Исследование безрецидивной и общей выживаемости пациентов в свете генетических подтипов позволяет сделать вывод о том, что прогностически более неблагоприятными вариантами можно считать N1 и MCD варианты (рис. 7).
Рисунок 7. Кривые безрецидивной (а) и общей (б) выживаемости больных ДВККЛ различных генетических подтипов.
Рисунок 8. Интегральная схема, отражающая взаимоотношение молекулярных подтипов лимфомы на основе профиля экспрессии генов и генетических подтипов, а также потенциальные молекулярные мишени для терапевтических приложений для того или иного генетического типа.
Несмотря на очевидный прогресс в понимании молекулярно-генетического разнообразия и патогенеза ДВККЛ и вопреки многообещающим результатам, полученным при исследовании токсического эффекта таргетных лекарственных препаратов на опухолевые клетки в экспериментальных моделях, на текущий момент в самых последних обновлениях американских клинических онкологических рекомендаций NCCN и в европейских рекомендациях медицинской онкологии ESMO альтернатив режиму R-CHOP в первой линии терапии нет [13].
Такие несоответствия обусловлены тем, что не всегда логичная и стройная идея об эффективности лекарственного препарата, подтвердившаяся в доклинических исследованиях, сможет оправдать себя в клинических исследованиях, в которых принимают участие живые больные люди. Более того, даже наличие статистически значимых различий между экспериментальным лечением и золотым стандартом или плацебо в пользу экспериментального лекарственного препарата не говорит о реальной клинической значимости, выраженной в количестве месяцев или лет жизни, прожитых пациентом в ремиссии в хорошем качестве. Уже имеющиеся результаты крупных клинических исследований II и III фаз по лечению пациентов с впервые выявленной ДВККЛ в попытке улучшить результаты лечения путем добавления ибрутиниба (исследование PHOENIX), леналидомида (исследования ROBUST и E1412) или бортезомиба (исследование REMoDL-B) к стандартному режиму терапии R-CHOP в первой линии, к сожалению, не увенчались успехом [14–17].
Вместо заключения. Зачем изучать такую сложную молекулярную биологию рака?
Если вы уже устали, дойдя до этого момента, из-за всех сложных биологических концепций разнообразия ДВККЛ, то я поспешу вас обрадовать тем, что в этом обзоре освещается лишь только один гистологический вариант лимфомы. Да, современные молекулярно-биологические представления опухолевого роста разнообразны и огромны. Одних внутриклеточных молекулярных посредников, участвующих в каскадных сигналах, существует сколько! А сколько мутаций во всех раковых генах! Весь этот массив информации в голове просто не удержать.
Сегодняшний рынок онкологического биофармацевтического производства представляет совокупность лекарственных препаратов, назначение которых основано на знаниях генетики рака, создавая тем самым возможность прицельного, направленного воздействия на опухоль. Да, да, вы, наверное, уже догадались, что я сейчас говорю именно о персонифицированной или персонализированной медицине в онкологии. Клиницисты сегодня знают довольно немало таких примеров: назначение эрлотиниба пациенту с аденокарциномой легкого при наличии мутации L858R в гене рецептора эпидермального фактора роста EGFR, вемурафениб для лечения распространенной меланомы при обнаруженной мутации в опухоли V600E в гене BRAF, иматиниб как блокатор BCR-ABL при терапии хронического миелолейкоза, мидостаурин в лечении острого миелоидного лейкоза при выявленной мутации в FLT3. Всего этого и многого чего другого просто не было, если бы не развивалась биомедицинская наука. От знаний в молекулярной биологии и генетики выигрывают абсолютно все: ученые удовлетворяют собственный научный интерес и любопытство, врачи принимают участие в клинических исследованиях, развивается фармацевтический рынок, появляются новые профессиональные кадры, а пациенты дольше живут, имея хорошее качество жизни.
Пусть сегодня успехи в выборе клинически эффективного таргетного лекарственного препарата на основе молекулярно-генетических характеристик ДВККЛ не так грандиозны, как хотелось бы, совершенно очевидно одно — дальнейшее усложнение наших представлений в отношении молекулярной генетики опухолей, особенно в условиях расширенной доступности технологий высокопроизводительного секвенирования непременно приведут к революционным терапевтическим подходам в не столь отдаленном будущем. В заключение стоит отметить, что попытка модифицировать схему R-CHOP предпринимается сейчас авторами исследования POLARIX, где они предлагают сравнить режим Pola-R-CHP (полатузумаб ведотин, ритуксимаб, циклофосфамид, доксорубицин, преднизолон) со стандартом, и уже получены результаты: 2-летняя выживаемость без прогрессирования у пациентов из группы лечения Pola-R-CHP составила 76,7% в сравнении с 70,2% у пациентов стандартной терапии R-CHOP. Более того, за последние два года появились возможности лечения рефрактерных или рецидивирующих форм ДВККЛ, такие как биспецифические моноклональные антитела (глофитамаб, мосунетузумаб, эпкоритимаб, одронекстамаб) и CAR-T клеточная терапия.
Читайте также: