
Вирусная теория канцерогенеза по
Обновлено: 18.04.2024
Согласно теории Вирхова, патология клетки лежит в основе любой болезни. Канцерогенез - последовательный, многоступенчатый процесс накопления клеткой изменений ключевых функций и характеристик, приводящих к ее озлокачествлению. Клеточные изменения включают нарушения регуляции процессов пролиферации, дифференцировки, апоптоза и морфогенетических реакций. В результате клетка приобретает новые качества: иммортализацию ("бессмертие", т.е. способность к неограниченному делению), отсутствие контактного ингибирования и способность к инвазивному росту. Кроме того, опухолевые клетки получают способность избегать действия факторов специфического и неспецифического противоопухолевого иммунитета организма хозяина. В настоящее время ведущая роль в индукции и промоции канцерогенеза принадлежит генетическим нарушениям. Около 1% генов человека ассоциированы с канцерогенезом.
4 стадии канцерогенеза:
1. Стадия инициации (изменение клеточных онкогенов, выключение генов-супрессоров)
· Фаза метаболической активации( превращение проканцерогенов в канцерогены)
· Фаза взаимодействия с ДНК ( прямой и непрямой генотоксический эффект)
· Фаза фиксации индуцированных изменений (повреждения ДНК должны проявиться в потомстве клеток-мишеней, способных давать пролиферативный пул.)
2. Стадия промоции
I(ранняя) фаза- перестройка фенотипа, происходящая вследствие эпигенетических изменений (т.е. генной экспрессии), индуцированных опухолевым промотором.
Изменение генной экспрессии, что дает возможность клетке функционировать в условиях пониженного синтеза генных продуктов.
II (поздняя) фаза - представляет собой качественно-количественные изменения, охватывающие период функционирования клетки в условиях переключения генной активности, завершающийся образованием неопластически трансформированных клеток (неопластическая трансформация — проявление признаков, характеризующих возможность клеток к неограниченной пролиферации и дальнейшей профессии, т.е. накоплению злокачественного потенциала
3. Стадия прогрессии: разработана L.Foulds в 1969 г. Происходит постоянный стадийный прогрессирующий рост опухоли с прохождением ею ряда качественно отличных стадий в сторону повышения ее злокачественности. В ходе прогрессии опухоли может происходить ее клональная эволюция, появляются новые клоны опухолевых клеток, возникающие в результате вторичных мутаций. Опухоль постоянно изменяется: происходит прогрессия, как правило, в сторону повышения ее злокачественности, которая проявляеются инвазивным ростом и развитием метастазов.
Стадияинвазивной опухолихарактеризуется возникновением инфильтрирующего роста. В опухоли появляются развитая сосудистая сеть и строма, выраженная в различной степени. Границы с прилежащей неопухолевой тканью отсутствуют из-за прорастания в нее опухолевых клеток. Инвазия опухоли протекает в три фазы и обеспечивается определенными генетическими перестройками.
Первая фаза инвазии опухоли характеризуется ослаблением контактов между клетками, о чем свидетельствуют уменьшение количества межклеточных контактов, снижение концентрации некоторых адгезивных молекул из семейства CD44 и других и, наоборот, усиление экспрессии прочих, обеспечивающих мобильность опухолевых клеток и их контакт с экстрацеллюлярным матриксом. На клеточной поверхности снижается концентрация ионов кальция, что приводит к повышению отрицательного заряда опухолевых клеток. Усиливается экспрессия интегриновых рецепторов, обеспечивающих прикрепление клетки к компонентам экстрацеллюлярного матрикса - ламинину, фибронектину, коллагенам.
Во второй фазе опухолевая клетка секретирует протеолитические ферменты и их активаторы, которые обеспечивают деградацию экстрацеллюлярного матрикса, освобождая тем самым ей путь для инвазии. В то же время продукты деградации фибронектина и ламинина являются хемоаттрактантами для опухолевых клеток, которые мигрируют в зону деградации в ходе третьей фазыинвазии, а затем процесс повторяется снова.

4. Стадия метастазирования - заключительная стадия морфогенеза опухоли, сопровождающаяся определенными гено- и фенотипическими перестройками опухоли. Процесс метастазирования связан с распространением опухолевых клеток из первичной опухоли в другие органы по лимфатическим и кровеносным сосудам, периневрально, имплантационно, что стало основой выделения видов метастазирования. Процесс метастазирования объясняется теорией метастатического каскада, в соответствии с которой опухолевая клетка претерпевает цепь (каскад) перестроек, обеспечивающих распространение в отдаленные органы.
В процессе метастазирования опухолевая клетка должна обладать качествами:
· проникать в прилежащие ткани и просветы сосудов (мелких вен и лимфатических сосудов);
· отделяться от опухолевого пласта в ток крови (лимфы) в виде отдельных клеток или небольших их групп;
· сохранять жизнеспособность после контакта в токе крови (лимфы) со специфическими и неспецифическими факторами иммунной защиты;
· мигрировать в венулы (лимфатические сосуды) и прикрепляться к их эндотелию в определенных органах;
· инвазировать микрососуды и расти на новом месте в новом окружении.
Метастатический каскад условно может быть разделен на четыре этапа:
1. формирование метастатического опухолевого субклона;
2. инвазия в просвет сосуда;
3. циркуляция опухолевого эмбола в кровотоке (лимфотоке);
4. оседание на новом месте с формированием вторичной опухоли.
В настоящее время существует несколько концепций онкогенеза , каждая из которых преимущественно влияет на 1 и(или) 2 этап канцерогенеза
Мутационная теория канцерогенеза
Нормальная клетка превращается в опухолевую в результатеструктурных изменений в генетическом материале, т.е. мутаций. Стало аксиомой представление о многоэтапности процесса канцерогенеза, решающей предпосылкой которого является нерегулируемая экспрессия трансформирующего гена – онкогена, предсуществующего в геноме.
Превращение протоонкогена в активно действующий онкоген обеспечивается следующими механизмами.
1. Присоединение к протоонокгену промотора– участка ДНК, с которым связывается РНК-полимераза, инициирующая транскрипцию гена, в том числе и онкогена, располагающегося непосредственно за ним . Такого рода участки (промоторы) содержатся в больших терминальных повторах (LTR)ДНК-копий РНК-содержащих вирусов. Роль промотора могут выполнять и транспозирующие элементы генома – мобильные генетические элементы, способные перемещаться по геному и встраиваться в различные его участки
2. Вставка в геном клетки энхансера(enchancer – усилитель) – участка ДНК, способного активизировать работу структурного гена, находящегося не только в непосредственной близости от него, но и на расстоянии многих тысяч пар нуклеотидов или даже встроенного в хромосому после него. Свойствами усилителя обладают подвижные гены, LTR ДНК-копий.
3. Хромосомные абберации с явлениями транслокации,роль которых в механизмах опухолевой трансформации клетки можно проиллюстрировать следующим примером.
При лимфоме Беркитта конец q-плеча хромосомы 8, отделившись от нее, переходит к хромосоме 14: гомологичный фрагмент последней перемещается к хромосоме 8; а неактивный гентуc (протоонкоген), находившийся в том ее сегменте, который попадает на хромосому 14, встраивается вслед за активными генами, кодирующими тяжелые цепи молекул иммуноглобулинов, и активизируется . Явления реципрокной транслокации между 9-й и 22-й хромосомами имеют место в 95 % случаев миелоцитарного лейкоза. Хромосома 22 с укороченным в результате такой транслокации одним плечом получила название Филадельфийской.
4.Точечные мутации протоонкогена,к примеру, C-H-raS, согласно имеющимся сведениям, отличается от нормального гена (C-H-raS) всего одной аминокислотой, но, тем не менее обусловливает снижение гуанозинтрифосфатазной активности в клетке, что может вызвать рак мочевого пузыря у человека.
· могут модифицировать рецепторы факторов роста
· действовать на ключевые внутриклеточные процессы
Тканевая теория канцерогенеза
Клетка становится автономной, т.к. нарушается тканевая система контроля пролиферации клоногенных клеток, обладающих активизированными онкогенами. Основным фактом, подтверждающим механизм, основанный на нарушении тканевого гомеостаза, является способность опухолевых клеток нормализоваться при дифференцировке.Изучение перевивной ороговевающей карциномы крысы методом автографического анализа показало (Pierce, Wallace, 1971), что раковые клетки при делении могут давать нормальное потомство, то есть злокачественность генетически не закреплена и не наследуется дочерними клетками, как это предполагалось мутационной гипотезой и молекулярно-генетической теорией. Хорошо известны эксперименты по пересадке ядер опухолевых клеток в предварительно энуклеированные зародышевые клетки: в этом случае развивается здоровый мозаичный организм. Таким образом, вопреки представлению о якобы сохранении трансформированных онкогенов в нормализованных опухолевых клетках при дифференцировке, есть основание поставить под сомнение связь генетических нарушений с механизмом трансформации в качестве непосредственной причины.
Вирусная теория канцерогенеза
Основателем вирусной онкологии следует считать Пэйтона Рауса, который в 1911 г. описал куриную саркому, перевиваемую от птицы к птице с помощью бесклеточных фильтратов. Вирус получил название вируса саркомы Рауса.
Чтобы стать злокачественной клетка должна приобрести по крайней мере 6 свойств как результат мутации генов, ответственных за деление клетки, апоптоз, репарацию ДНК, внутриклеточные контакты и т.д. В частности, на пути к приобретению злокачественности клетка, как правило:
1) самодостаточна в плане сигналов пролиферации (что может быть достигнуто активацией некоторых онкогенов, например, Н-Ras);
2) нечувствительна к сигналам, подавляющим ее рост (что происходит при инактивации гена опухолевого супрессора Rb);
3) способна ослабить или избежать апоптоза (что происходит в результате активации генов, кодирующих факторы роста);
4) формирование опухоли сопровождается усиленным ангиогенезом (что может быть обеспечено активацией гена VEGF, кодирующего ростовые факторы эндотелия сосудов;
5) генетически нестабильна;
6) не подвергается клеточной дифференцировке;
7) не подвергается старению;
8) характеризуется изменением морфологии и локомоции, что сопровождается приобретением свойств к инвазии и метастазированию.
Поскольку мутации генов являются событиями случайными и достаточно редкими, их накопление для инициации клеточной трансформации может длиться десятилетиями. Трансформация клетки может произойти гораздо быстрее в случае высокой мутагенной нагрузки и/или дефектности (слабости) механизмов защиты генома (генов p53, Rb, ДНК репарации и некоторых других). В случае же инфицирования клетки онкогенными вирусами, кодируемые вирусным геномом белки, обладающие трансформирующим потенциалом, нарушают номальные клеточные сигнальные связи, обеспечивая условия для активной клеточной пролиферации.
Хорошо известно, что возникновение примерно 15-20% новообразований человека имеют вирусное происхождение. Среди наиболее часто встречающихся таких вирусом индуцированных опухолей можно назвать рак печени, рак шейки матки, рак носоглотки, лимфому Беркитта, лимфому Ходжкина и многие другие. В настоящее время экспертами Международного Агентства по Изучению Рака (МАИР) следующие вирусы рассматриваются в качестве онкогенных для человека:
Вирусы гепатита В и С (Hepatitis B virus и Hepatitis C virus, HBV/ HCV), вызывающие рак печени; В результате генетических перестроек происходит делеция гена X и некоторой части генов PreS2, при этом клетки печени становятся HBsAg-негативными и окончательно уходят из-под иммунологического контроля. Далее происходит селекция клеток, в которых интегрирована ДНК HBV и которые содержат
3 основных транс-активатора, а именно: HBx, LHBs и/или MHBs(t). Транс-активаторы активируют клеточные гены, ответственные за пролиферацию клеток, синтез цитокинов (IL-6) и т.д. Цитокины, секретируемые клетками, содержащими транс-активаторы, создают микроокружение из прилегающих фибробластов, эндотелиальных клеток и др., в свою очередь, выделяющих другие ростовые факторы, стимулирупующие по паракринному типу пролиферацию гепатоцитов. Усиленная пролиферация гепатоцитов может привести к генетическим поломкам, которые будут способствовать селекции клеток с ускоренной пролиферацией и приобретению ими признаков злокачественной трансформации. В опухолевых клетках печени часто имеет место инактивация опухолевых супрессоров р53, Rb, BRCA2 и Е-кадхерина. Отмечена также активация теломеразы в печеночных клетках на стадии их превращения в злокачественные и нарушение функционирования ряда важных сигнальных систем.
Определенные типы (16 и 18) папаломавирусов человека (Human papillomavirus, HPV) — являющихся этиологическим агентом рака шейки матки и некоторых опухолей ано-генитальной сферы; Установлено, что трансформирующими генами являются в основном гены Е6 и Е7, в меньшей степени Е5. Механизм функционирования генов Е6 и Е7 сводится к взаимодействию продуктов этих генов с продуктами 2-х генов супрессоров р53 и Rb и последующей инактивации последних, что приводит к неконтролируемому росту инфицированных клеток.Проведенные исследования показали, что каждый из выше упомянутых 3-х генов латентной HPV инфекции, обладающий трансформирующими потенциями, вносит свой вклад внарушение сигнальных путей клетки, увеличение ее пролиферативной активности и накопление дополнительных генетических изменений. Стоит отметить что созданы терапевтические и профилактические вакцины против ВПЧ. Которые стимулюруют иммунную систему против Е6 и/или Е7 ранних вирусных белков (опухолевых антигенов), препятствующих входу инфицированных клеток в апоптоз и фазу старения, а также генерируют вирус-нейтрализующие антитела, специфические для капсида HPV.
Вирус Эпштейна-Барр (Epstein-Barr virus, EBV), принимающий участие в возникновении целого ряда злокачественных новообразований;Механизм канцерогенеза сложен и мало изучен. В частности, белок LMP1, локализуясь в мембране, имитирует функцию конститутивно активированного рецептора СD40 и частично замещает эту функцию. Привлекая адаптерные молекулы TRAF через домены активации CTAR1 и CTAR2 активирует транскрипционные факторы AP-1 и NFkB и таким образом индуцирует экспрессию генов, регулируемую этими факторами (рецептор эпидермального фактора роста, EGFR, CD40, поверхностные активационные маркеры, молекулы адгезии и т.д.). Кроме того, LMP1 взаимодействует с Jak3-киназой и таким образом активирует STAT-сигнальные пути, стимулирующие размножение и передвижение клеток. LMP2A активирует киназу Akt/PBK, вызывая ряд эффектов, наиболее ярким из которых является подавление апоптоза. EBNA2 имитирует транскрипционную функцию процессированной формы Notch (трансмембранный белок, преобразующий контакты с окружающими клетками в генетические программы, регулирующие судьбу клетки), конститутивная активность которого ведет к развитию лимфоидных и эпителиальных опухолей. Основная функция EBNA1 состоит в обеспечении репликации и поддержания эписомального состояния генома ВЭБ.
Вирус Т-клеточного лейкоза человека (Human T-cell leukemia virus, HTLV-1), являющийся этиологическим агентом Т-клеточного лейкоза взрослых, а также тропического спастического парапареза и ряда других неонкологических заболеваний.Механизм транс-актививации транскрипции ряда вирусных и клеточных генов (цитокинов, их рецепторов, циклинов и др), ассоциированных с клеточной пролиферацией и способствующих росту инфицированных HTLV-1 клеток. Белок Тах может и транс-репрессировать транскрипции определенных генов, действуя через транскрипционный ко-активатор р300. Тах также инактивирует чекпоинты (сверочные точки) клеточного цикла и ДНК-полимеразу (DNApol), снижая активность всех 3-х систем репарации ДНК и вызывая тем самым генетическую нестабильность, что в конечном итоге приводит к возникновению опухолевой клетки.
Вирус иммунодефицита человека (Human immunodeficiency virus, HIV)— не обладающего трансформирующими генами, но создающего необходимые условия (иммунодефицит) для возникновения рака.
Несмотря на различную организацию онкогенных вирусов человека, неодинаковый спектр их клеток-мишеней, они обладают рядом общих биологических свойств, а именно:
1) вирусы лишь инициируют патологический процесс, усиливая пролиферацию и генетическую нестабильность инфицированных ими клеток;
2) у инфицированных онкогенными вирусами лиц возникновение опухоли, как правило, событие нечастое: один случай новообразования возникает среди сотен, иногда тысяч инфицированных;
3) после инфицирования до возникновения опухоли имеет место продолжительный латентный период, длящийся годами, иногда десятилетиями;
4) у большинства инфицированных лиц возникновение опухоли не является обязательным, но они могут составить группу риска, с более высокой возможностью ее возникновения;
5) для злокачественной трансформации инфицированных клеток необходимы дополнительные факторы и условия, приводящие к селекции наиболее агрессивного опухолевого клона.
ДНК-вирусы как причина развития рака - вирусный канцерогенез
В канцерогенезе также могут участвовать ДНК-содержащие вирусы. Наиболее крупные ДНК-содержащие вирусы принадлежат к группе герпесвирусов. Первым вирусом, выделенным из культуры опухолевых клеток, был вирус Эпштейна-Баррра (ЭБВ).
Вирус был получен из культуры опухолевых клеток больных лимфомой Беркита. В культуре in vitro ЭБВ вызывает бесконечную пролиферацию человеческих В-лимфоцитов. Вирусные гены, встраиваясь в геном этих клеток, инициируют пролиферацию путем активации клеточных генов-регуляторов роста (онкогенов).
В крови больных лимфомой Беркита обнаруживается более высокий, чем у здоровых людей, титр антител к антигенам вирусной оболочки и к мембранным антигенам. В то же время в африканских странах среди множества детей, инфицированных этим вирусом, лимфома развивается только в единичных случаях.
Механизм онкогенеза этого заболевания не может быть объяснен только вирусным заражением. К тому же, за пределами африканского континента регистрируются случаи заболевания лимфомой Беркита, абсолютно не связанные с инфицированием вирусом ЭБВ. По-видимому, ЭБВ также может принимать участие в патогенезе карциномы носоглотки, так как клетки этой опухоли экспрессируют вирусные антигены EBNA-1.
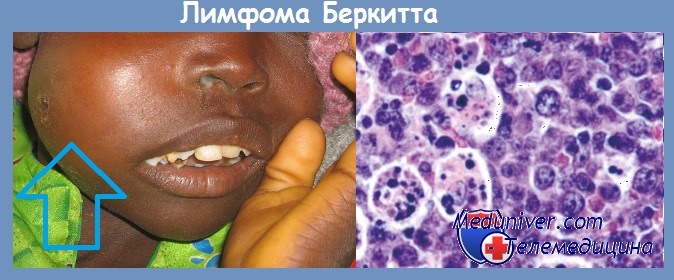
При исследованиях саркомы Капоши, развивающейся у больных СПИДом, в 1994 г. из ее клеток был выделен новый герпесвирус. Первоначально вирус получил название герпес-вируса саркомы Капоши (ГСК), впоследствии он был переименован в герпесвирус человека серотип 8 (ГВЧ 8). Вирус обнаруживается в веретеновидных клетках саркомы Капоши практически при всех случаях заболевания.
ГВЧ 8 был также найден в опухолевых клетках первичной эффузионной лимфомы и в клетках многоочаговой болезни Кастлемана. Данный вирус кодирует синтез циклина, который является промотором клеточного деления, чем и объясняется высокая пролиферативная активность инфицированных клеток.
Человеческие папилломавирусы (ПВЧ) вызывают кожные бородавки и ряд доброкачественных папиллом. Это весьма обширный класс вирусов, и некоторые его представители могут в той или иной степени участвовать в развитии злокачественных опухолей. Большинство вирусов вызывает лишь ограниченную пролиферацию клеток, но ряд серотипов может вызвать и развитие злокачественных новообразований.
Наибольшую опасность с точки зрения онкологии представляют серотипы 16, 18, 31 и 33. Более 90% опухолей карциномы шейки матки in situ обнаруживают в своих клетках генные последовательности вирусов ПВЧ. Эти вирусы также играют значительную роль в возникновении инвазивных и многоочаговых плоскоклеточных раков кожи, которые часто развиваются у больных после долгосрочных курсов иммуносупрессивной терапии.
Механизмы вирусного канцерогенеза в этом случае изучены недостаточно. В исследованиях доказано, что существует вероятность включения вирусной ДНК в клетки базального эпителия. Обычно в клеточную ДНК внедряются восемь ранних и два поздних гена. Ранние гены Е6 и Е7 вызывают пролиферацию клеток.
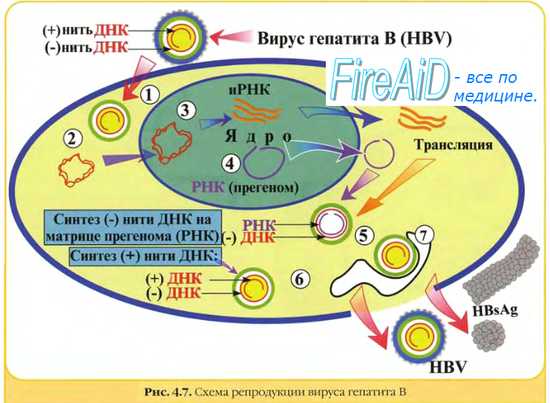
Вирус гепатита В четко связывают с развитием печеночно-клеточного рака. Риск развития этого заболевания у инфицированных вирусом в 200 раз превышает таковой у неинфицированных людей. Механизмы зарождения рака не до конца понятны: считается, что вирус вызывает бесконечную пролиферацию клеток, хотя зараженные вирусом клетки уничтожаются иммунной системой организма. Возможно, что пролиферирующие клетки становятся чувствительными к другим канцерогенам, например к афлатоксину.
Подытоживая вышесказанное, можно заключить, что основным механизмом канцерогенного воздействия является повреждение молекул ДНК и нарушения процессов ее репарации. Если не происходит исправления первичных повреждений ДНК, ошибки со временем начинают накапливаться, что ведет к дестабилизации структуры хромосом и к соматическим мутациям. Большинство таких мутаций летальны для клетки.
Однако, если часть клеток выживает, происходит постепенное накопление клеточной массы с опухолевым фенотипом, что в итоге приводит к клиническим проявлениям. Большой вклад в понимание этого процесса внесли современные исследования механизмов регулировки клеточного цикла и недавнее открытие регуляторных белков, которые запрещают клетке делиться, если ее геном сильно поврежден.
Для развития опухоли может быть недостаточно единичных мутаций в генах-регуляторах деления, но накопление массы ошибок в регуляторных генах может привести к фатальным последствиям. Особенно критичны мутации в генах, регулирующих клеточное деление, так называемых онкогенах. Очень опасны также мутационная инактивация генов, подавляющих клеточное деление — генов-подавителей опухоли, и мутации в генах, кодирующих белки, запрещающие деление при повреждении клеточного генотипа.
Накопление числа жизнеспособных мутаций по одному или нескольким из этих генов приводит к общей клеточной дезорганизации и к потере контроля над клеточным делением. В следующем разделе мы рассмотрим механизмы действия онкогенов и наиболее важные моменты контроля над клеточным циклом.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
РНК-вирусы как причина развития рака - вирусный канцерогенез
Открытие вирусов, вызывающих злокачественные опухоли у животных, произошло еще на рубеже XIX и XX веков. В 1910 г. Пейтон Рауш обнаружил, что бесклеточный фильтрат из тканей птичьей саркомы может вызывать развитие аналогичной саркомы у цыплят. Примерно в это же время была доказана вирусная природа птичьего миелобластоза. Позднее было обнаружено, что часто проходит весьма значительный латентный период между инфицированием вирусом и развитием рака.
Тем не менее вплоть до 1960-х годов не существовало четких доказательств того, что инкорпорация вирусной ДНК в геном клетки является необходимым условием развития злокачественной трансформации, как не было и случаев выделения вирусной ДНК из раковых клеток.
РНК-содержащие вирусы вызывают развитие целого ряда различных опухолей у животных, при этом наиболее распространена индукция этими вирусами лимфом, лейкемий и сарком. Типичная структура таких вирусов - это две идентичные цепочки молекул РНК в сочетании с ферментом обратная транскриптаза, одетых в гликопротеиновую оболочку. При инфицировании вирусом его обратная транскриптаза заставляет клетки синтезировать ДНК, комплементарную вирусной РНК.
Эта ДНК затем встраивается в клеточные хромосомы, и на ее основе клетка уже сама начинает синтез новых вирусных белков, вирусных обратных транскриптаз и элементов гликопротеиновой оболочки. Из-за механизма их действия этот тип вирусов получил название ретровирусы. Все они имеют весьма схожий внешний вид на электронных микрофотографиях и являются самыми маленькими из известных вирусов.
Некоторые из ретровирусов (например, вирусы птичьего лейкоза, кошачьей и мышиной лейкемии) содержат в себе только три гена и обладают очень длительным инкубационным периодом с момента заражения до возникновения опухоли. Другие вирусы (например, вирус саркомы Рауса (ВСР)) вызывают очень быструю злокачественную трансформацию и могут быть выделены из культуры опухолевых клеток.
Показано, что вирус ВСР содержит особый ген (v-src), способный вызывать трансформацию фибробластов in vitro. Этот ген кодирует наработку протеинкиназы, которая фосфорилирует тирозин. К сожалению, действие этой протеинкиназы запускает целый каскад различных метаболических процессов, и очень сложно оценить, какой именно из них ведет к злокачественной трансформации.
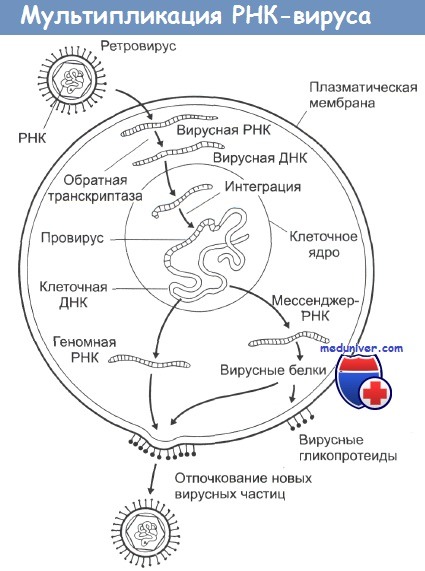
В настоящее время известно, что и нормальные, и злокачественные клетки содержат в своем генотипе участки ДНК, сходные или идентичные ряду последовательностей онкогенных РНК-содержащих вирусов. Такие участки получили название клеточных прото-онкогенов (чтобы отличать их от вирусных онкогенов). Постулируется, что активация этих участков, возникающая в результате канцерогенного воздействия, запускает целую цепочку событий, ведущую в итоге к злокачественной трансформации клетки. Считается также, что ретровирусы инкорпорировали эти клеточные участки в свой геном в процессе эволюции.
В настоящее время мы лучше понимаем механизмы действия продуктов вирусной активации. Примером такого продукта является протеинкиназа, активируемая геном sre, а также целый ряд других вирус-индуцированных канцерогенов. Это и рецепторы к эпидермальному фактору роста, продуцируемые геном v-erb, и тромбоцитарный фактор роста (ТФР), кодируемый фрагментами гена v-sis, и целый ряд связывающихся с ядром клетки белков, продукцию которых вызывает вирус птичьей лейкемии.
Как нормальные, так и раковые клетки содержат в себе участки последовательностей ДНК, гомологичные РНК онкогенных вирусов. Если происходит экспрессия или активация под действием канцерогенов этих клеточных онкогенов, это ведет к злокачественной трансформации клеток.
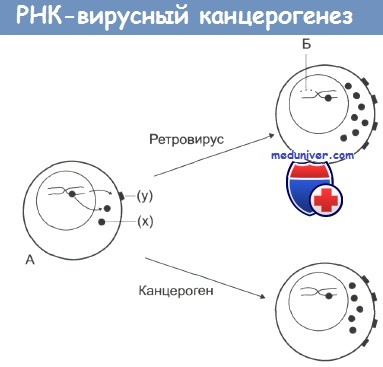
Онкогенная и злокачественная трансформация.
На стадии А нормальная клетка, которая характеризуется низкой протоонкогенной активностью, продуцирует фактор роста (х) или белки дифференцировки или рецепторы (у).
Канцерогены повышают активность протоонкогенов, что дает начало неопластической трансформации.
По другому механизму: при заражении ретровирусом происходит внедрение в ДНК клетки вирусных промоторов или онкогенов (Б), что также ведет к увеличению онкогенной активности и последующей злокачественной трансформации.
Первым ретровирусом, для которого была однозначно доказана связь со злокачественными новообразованиями, был вирус Т-клеточной лейкемии человека (ВТКЛ-1), выделенный из клеток хронической кожной Т-лимфомы. Этот вирус довольно широко распространен, может передаваться половым путем, через кровь, особенно в среде наркоманов, и от беременной женщины к плоду. Первоначально эндемический ареал циркуляции данного вируса был в основном представлен тропическими странами, однако в настоящее время в США серопозитивная реакция на вирус обнаруживается у каждого из 4000 человек населения. Кроме Т-клеточной лейкемии вирус вызывает тропический спастический паралич.
После 20 лет наблюдения за серопозитивными пациентами оценено, что риск развития последнего заболевания составляет около 5%. Один из вирусных генов, а именно tax-ген, вызывает увеличение продукции клеточного интерлейкина-2 (ИЛ-2) и его рецепторов, что является основным фактором, стимулирующим деление Т-клеток.
Ретровирусы могут вызывать опухолевые заболевания не напрямую, а опосредованно, как это было показано для вируса иммунодефицита человека (ВИЧ-1), вызывающего развитие СПИДа. Случаи развития рака у ВИЧ-инфицированных рассмотрены в одном из обзоров. Отмечается, что у ВИЧ-инфицированных наиболее часто развиваются три типа опухолей: мгновенная или высокозлокачественная В-клеточная лимфома; саркома Капоши (СК, которую вызывает другой вирус — герпесвирус ГСК, или герпесвирус 8); карцинома шейки матки.
До разработки эффективных методов терапии ВИЧ-инфицированных более 40% из них заболевали тем или иным видом рака. Тем не менее связь этого вируса с развитием онкологических заболеваний скорее всего косвенная и опосредована развитием общей хронической иммунодепрессии организма, которая и позволяет другим канцерогенным вирусам вызывать рак. В-клеточные лимфомы также характеризуются довольно сложным патогенезом. Хотя В-клетки не поражаются вирусом ВИЧ-1, они могут быть мишенью для других типов вирусов, например для вируса Эпштейна-Барра (ВЭБ). Рак шейки матки у женщин также развивается на основе вторичной вирусной инфекции — вируса папилломы человека (ВПЧ), на фоне общей иммунодепрессии, вызванной ВИЧ-1. По причине общего снижения иммунных реакций организма все эти опухоли развиваются особенно быстро и агрессивно.
Показано, что РНК-содержащий вирус гепатита С значительно увеличивает риск заболевания гепатоклеточной карциномой. У больных, инфицированных этим вирусом, риск развития раковой опухоли возрастает в 100 раз, но механизмы его канцерогенного действия в настоящее время почти не изучены. Если больной заражен еще и вирусом гепатита В, риск развития рака печени возрастает многократно. На земном шаре одним или обоими из этих вирусов заражено около миллиарда человек.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Вирусная теория развития рака. Вирусный канцерогенез.
В 1908—1911 гг была установлена вирусная природа лейкоза и саркомы кур. В последующие десятилетия была доказана вирусная этиология ряда лимфоидных и эпителиальных опухолей у птиц и млекопитающих. В настоящее время известно, что в естественных условиях, например, лейкоз вызывается вирусами у цыплят, кошек, крупного рогатого скота, мышей, обезьян-гиббонов.
В последние годы открыт первый вирусный возбудитель, вызывающий развитие лейкоза у человека,— это ATLV (adult T-cell leukemia virus — вирус Т-клеточного лейкоза взрослых) Т-клеточный лейкоз взрослых — эндемичное заболевание, встречающееся в двух районах земного шара иа островах Клуши и Шихоку в Японском море и у негритянского населения стран Карибского бассейна. Больные с этой лимфомой встречаются спорадически н в других регионах, но у многих из них выявлена та или иная связь с эндемическими областями.
Заболевание это встречается обычно у людей старше 50 лет, протекает с кожными поражениями, гепатомегалией, спленомегалией, лимфаденопатией и имеет неблагоприятный прогноз Вирус ATLV или HTLV является экзогенным для человека, отличается от других известных ретровирусов животных, передается Т-клеткам горизонтально от матери к ребенку, от мужа к жене (но не наоборот), при лереливании крови, не обнаруживается ни при каких других формах лейкозов или лимфом человека. Таким образом, Т-клеточный лейкоз взрослых — это типичное инфекционное заболевание (вертикальная передача вируса через половые клетки исключена специальными исследованиями). В эндемичных очагах более 20% практически здоровых людей, главным образом родственники больных, являются носителями вируса.
В других частях земного шара антитела к вирусу обнаруживают редко. Считается, что заболевает 1 из 2000 инфицированных людей. Вирус, неотличимый от ATLV, обнаружен в Африке у обезьяи. Помимо лимфомы (лейкоза), указанный вирус может вызывать СПИД, при котором нарушен Т-клеточный иммунитет.
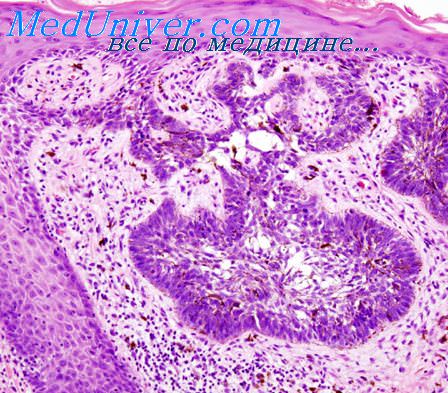
Вирусную этиологию подозревают и по отношению к некоторым другим опухолям человека Вирус Эпстайна — Барр (EBV), входящий в группу вирусов герпеса, является весьма вероятным этиологическим фактором лимфомы Беркитта. В клетках этой лимфомы в эндемических очагах в Африке постоянно обнаруживают ДНК EBV. Однако лимфома Беркитта встречается и за пределами Африки, но ДНК EBV обнаруживается лишь в меньшей части таких случаев. Общим для EBV-положительиых и EBV-отрицательных опухолей являются характерные перестройки хромосом (транслокация между хромосомами 8 и 14), что рассматривают как доказательство единой этиологии этих опухолей.
ДНК этого вируса находят в геноме клеток недифференцированной назофарингеальиой карциномы, но не в опухолях носоглотки другого гистогенеза. У больных с этими опухолями отмечают высокий титр антител к различным компонентам EBV, значительно превышающий эти показатели в популяции — EBV имеет широкое распространение, и антитела к нему обнаруживаются у 80—90% здоровых людей. Высокий титр антител обнаружен у больных лимфогранулематозом. Подавление иммунитета и активация EBV являются, по мнению ряда авторов, основной причиной развития лимфом и иммуиобластных сарком у больных с трансплантированными почками, подвергавшихся действию иммуноделрессивных средств; в пользу этого свидетельствует высокий титр антител к EBV и обнаружение ДНК EBV в геноме опухолевых клеток.
Существуют данные, позволяющие предполагать инфекционную (вирусную) этиологию рака шейки матки частота возникновения этого рака выше при раннем начале половой жизни с частой сменой партнеров, повышена она у вторых жен мужчин, первые жены которых также страдали тем же заболеванием. На основании сероэпидемиологических данных думают о роли вируса герпеса II типа как инициатора; подозревают также вирус кондилом.
В районах с высокой частотой возникновения вирусного гепатита В повышена также и заболеваемость гепатоцеллюлярным раком. С другой стороны, больные с этой опухолью чаще дают серопозитивную реакцию на вирус гепатита В, чем здоровые индивидуумы; но есть и серонегативные случаи рака. Получены линии опухолевых клеток, содержащие ДНК вируса и продуцирующие его антиген. В целом роль вируса гепатита В в индукции гепатоцеллюляриого рака остается невыясненной.
Из бородавок человека (verrucae vulgaris) выделено несколько типов вирусов папиллом, которые, как считают, вызывают лишь доброкачественные опухоли, не склонные к озлокачествлению. Лишь один из этих вирусов (5-го типа) выделен из папиллом, развивающихся при наследственной бородавчатой эпидермодисплазии и имеющих тенденцию к малигнизации.
Онкогены ныне идентифицированы, определена их химическая структура, локализация в хромосомах. Идентифицированы также белки — продукты деятельности этих генов, каждый из них синтезирует свой специфический белок.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Рак – бич человечества. По смертности он занимает второе место после сердечно-сосудистых заболеваний, по страху, который внушает людям – первое. Тысячи исследователей стремятся понять его причины, найти пути к его профилактике и лечению. Десятки институтов и сотни лабораторий во всем мире работают над этой проблемой, пытаясь достичь успеха в ее понимании и прогресса в профилактике и лечении этого заболевания.
Современной науке известны некоторые факторы, вызывающие развитие злокачественных опухолей и, вместе с тем, ученые вынуждены констатировать, что основная часть злокачественных опухолей возникает спонтанно, то есть без видимой связи с индуцирующими агентами. К факторам, способным вызывать развитие опухоли, относятся: различные канцерогенные вещества, наследственная предрасположенность, ионизирующее излучение и опухолеродные вирусы.
Вирусно-генетическая теория возникновения опухолей, предложенная еще в 40-х годах ХХ века Л.И. Зильбером, получила за прошедшие годы многочисленные подтверждения. В настоящее время, очевидно, что хотя вирусы и не являются единственной причиной рака, но они играют большую роль в возникновении злокачественных заболеваний как у человека, так и у животных. Характерной особенностью опухолевых заболеваний, ассоциированных с вирусами, является длительный латентный период, от момента инфекции до проявления заболевания могут пройти годы и даже десятилетия.
Согласно современным данным, этиологическими агентами около 15% опухолевых новообразований человека являются вирусы. К таким вирусам относятся: вирус Т-клеточного лейкоза/лимфомы (human T-leukemia/lymphoma virus), вирус иммунодефицита человека (ВИЧ), вирус папилломы человека (ВПЧ), вирусы гепатита В и С, вирус Эпштейна-Барр (ВЭБ) и другие. Важно отметить, что некоторые вирусы ассоциированы с опухолями только одной локализации, тогда как другие — с разными злокачественными новообразованиями, что, вероятно, обусловлено тропизмом вирусов к клеточным системам определенного типа.
Вирус Т-клеточного лейкоза/лимфомы (HTLV-1). HTLV-1 – это онкогенный вирус, способный вызывать Т-клеточный лейкоз/лимфому у взрослых, а также тропический спастический парапарез и ряд других неонкологических заболеваний.
Существуют 3 основных пути передачи инфекции от инфицированных лиц:
- с молоком матери;
- при половом контакте;
- с переливаемой кровью при гемотрансфузиях.
Основными методами диагностики вирусов гепатита В и С являются методы иммуноферментного анализа и полимеразной цепной реакции.
Вирус папилломы человека (ВПЧ, human papilloma virus). В середине 70-х гг. прошлого века было высказано предположение, что вирус папилломы человека является этиологическим агентом рака шейки матки, и эта область онкологии и вирусологии получила стремительное развитие. Действительно, к настоящему моменту роль ВПЧ в развитии рака шейки матки не вызывает сомнений. Многочисленные исследования доказали, что не менее 95% злокачественных опухолей шейки матки содержат разновидности ВПЧ, принадлежащие к так называемым типам "высокого онкогенного риска" (ВПЧ 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59 типов). Среди них максимальной онкогенной активностью обладают ВПЧ 16 и 18 типов. Действительно, ВПЧ достаточно часто (примерно в 30% случаев) выявляется у абсолютно здоровых женщин. Однако не у всех женщин, зараженных вирусом папилломы человека, возникает дисплазия шейки матки и рак шейки матки. Примерно у 80% женщин иммунная система организма в течение 2 лет после инфицирования сама избавляется от вируса. Таким образом, носительство этих вирусов свидетельствует не о злокачественном процессе как таковом, а о многократно повышенном риске его возникновения.
Диагностика ВПЧ-инфекции обладает очень высокой клинической значимостью. Она позволяет выявить тех пациенток, для которых жизненно необходимы активные, комплексные меры, направленные на профилактику и раннюю диагностику рака шейки матки. Самым надежным диагностическим тестом по выявлению всех 12 онкогенных типов вируса папилломы человека в настоящее время является метод полимеразной цепной реакции в режиме реального времени.
Вирус Эпштейна-Барр (ВЭБ). Известен широкий спектр патологических состояний, представленный опухолями лимфоидного и эпителиального происхождения, в возникновении которых ВЭБ принимает непосредственное участие. Доказано, что этот вирус причастен к возникновению не только лимфомы Беркитта, но и ряда других злокачественных и доброкачественных новообразований, таких, как рак носоглотки, лимфома Ходжкина, инфекционный мононуклеоз и многие другие.
Согласно некоторым исследованиям, ВЭБ широко циркулирует среди населения, о чем свидетельствует высокий процент людей, имеющих в крови антитела к ВЭБ - 50% детей и 85% взрослых. Но поскольку инфекция, как правило, протекает бессимптомно, сложилось ошибочное мнение о невысокой заболеваемости ВЭБ-инфекцией.
Вирус герпеса человека 8 типа (HHV-8). В настоящее время считается доказанной его роль в возникновении саркомы Капоши, выпотной лимфомы полостей тела и болезни Кастельмана. В геноме HHV-8 содержится ряд генов, продукты которых играют важную роль в регуляции размножения и жизнеспособности клеток; повышение их активности приводит к возникновению опухоли.
Следует понимать, что HHV-8 является всего лишь фактором риска развития вышеуказанных заболеваний. Если человек инфицирован HHV-8, то это не означает, что у него обязательно разовьётся саркома Капоши или лимфома.
Сам по себе вирус герпеса 8 типа опасности для здорового человека практически не представляет. Большинство людей с нормальным иммунитетом, инфицированных этим вирусом, не отмечают каких-либо проявлений, и он не оказывает на них никакого вредного влияния.
Свое негативное действие HHV-8 может проявить только в случае стойкого снижения иммунитета (иммунодепрессии), и то лишь у больных СПИДом (зараженных вирусом ВИЧ) или при длительном лечении, снижающем иммунитет (после пересадки органов и тканей, лучевой терапии). По данным статистики у 45% людей, имеющих HHV-8 и ВИЧ в течение 10 лет развилась саркома Капоши. У людей с нормальным иммунитетом вирус герпеса человека никак себя не проявляет. Он не влияет на состояние здоровья.
Таким образом, несмотря на различную организацию онкогенных вирусов человека они обладают рядом общих биологических свойств, а именно:
- вирусы лишь инициируют патологический процесс, усиливая процессы деления клеток и генетическую нестабильность инфицированных ими клеток;
- у инфицированных онкогенными вирусами лиц возникновение опухоли, как правило, событие нечастое: один случай новообразования возникает среди сотен, иногда тысяч инфицированных;
- от момента инфицирования до возникновения опухоли имеет место продолжительный латентный период, длящийся годами, иногда десятилетиями;
- у большинства инфицированных лиц возникновение опухоли не является обязательным, но они могут составить группу риска, с более высокой вероятностью ее возникновения;
- для злокачественной трансформации инфицированных клеток необходимы дополнительные факторы и условия, приводящие к прогрессированию наиболее агрессивного опухолевого клона.
Такими дополнительными факторами, играющими роль сопутствующих канцерогенов, могут быть: злоупотребление алкоголем, табакокурение, коинфекция генитальным герпесом, малярия, нитрозамины в продуктах питания, пестициды.
На сегодняшний день основными методами лабораторной диагностики являются метод иммуноферментного анализа (ИФА) и метод полимеразной цепной реакции (ПЦР). Первый позволяет выявить белок вируса либо антитела к нему, которые выделяет организм в ответ на инфицирование. При помощи метода ПЦР мы можем выявить генетический материал самого вируса.
Метод иммуноферментного анализа является стандартным методом лабораторной диагностики.
Следует также отметить, что не всегда возможно определение вирусных белков в плазме крови. В таких случаях методом ИФА можно диагностировать только наличие антител в плазме крови пациента. Однако, важно помнить, что антитела могут вырабатываться в довольно длительный срок. Так, например, при инфицировании ВИЧ антитела вырабатываются в течение 6 месяцев, в случае гепатита С этот период может длиться до года, а антитела к вирусу Эпштейна-Барр (иммуноглобулины класса G) присутствуют у большого количества практически здоровых людей и их диагностика обладает низкой прогностической значимостью.
Полимеразная цепная реакция (ПЦР) — метод молекулярной биологии, позволяющий обнаружить возбудителя заболевания на основе его генетической информации.
ПЦР диагностика дает возможность существенно ускорить и облегчить диагностику различных заболеваний, в частности, вирусов гепатита В, С, D, цитомегаловирусной инфекции, вируса Эпштейна-Барр, вируса простого герпеса, а также хламидиоза, микоплазмоза, уреаплазмоза и т.п. При помощи данного метода вирусные инфекции можно обнаружить уже через 5–7 дней после заражения, за недели или месяцы до того, как появятся антитела или первые симптомы. Метод ПЦР позволяет выявить даже единичные клетки возбудителя, благодаря многократному увеличению последовательностей ДНК.
Полимеразная цепная реакция - это один из самых точных методов лабораторной диагностики. Его чувствительность составляет 95–99%, а специфичность достигает 100%.
Таким образом, достижения науки в области онкологии внесли определенную ясность в понимание механизмов возникновения некоторых злокачественных новообразований и все же большую часть из них не удается увязать с инфицированностью тем или иным вирусом, что создает большое поле деятельности современным онкологам в направлении дальнейшего изучения механизмов развития онкологических заболеваний.
Все вышеуказанные методы диагностики существуют у нас в РНПЦ онкологии и медицинской онкологии им. Н.Н.Александрова. В лаборатории клинической молекулярной генетики и иммунологических методов диагностики можно пройти обследование на вирусы герпеса,цитомегаловируса, Эпштена-Барра, вирусы гепатита В и С, а также пройти обследование на инфицирование вирусом папилломы человека, данные исследования может пройти любой человек обратившийся в центр.
Читайте также: