
Вирусы использование в промышленности
Обновлено: 19.04.2024
Попадая в бактериальную клетку, фаги, как и положено вирусам, влияют на все процессы ее жизнедеятельности, фактически превращая клетку в фабрику по производству вирусного потомства. В конце концов клетка бактерии гибнет и разрушается, а вновь образованные вирусные частицы выходят наружу, чтобы найти себе новую жертву. А теперь представьте, что речь идет о бактерии, патогенной для человека или сельскохозяйственных животных и культурных растений. Как говорят, враги наших врагов… Даже в желудочно-кишечном тракте человека содержится около 10 12 вирусов-бактериофагов, а это говорит о многом.
Тем не менее базовые технологии генетической инженерии родились благодаря прикладным исследованиям в интересах молочной промышленности. Дело в том, что вирусные атаки на бактерий, использующихся при производстве сыра, йогурта и других кисломолочных продуктов, приносили ощутимые экономические потери, что потребовало создания устойчивых к бактериофагам штаммов.
На сегодня известны пять основных, весьма хитроумных механизмов защиты, которые либо препятствуют проникновению вирусной ДНК, либо уничтожают ее внутри бактериальной клетки.
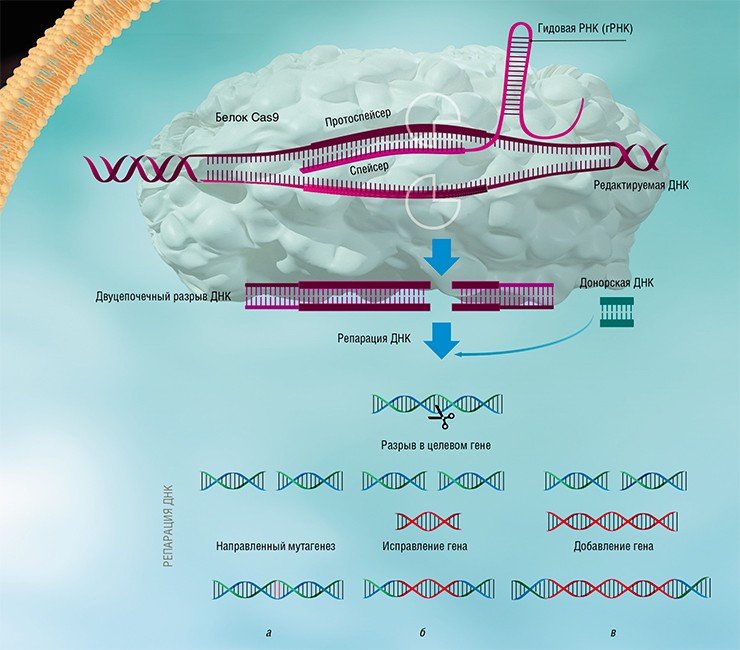
Все системы адаптивного иммунитета CRISPR-Cas действуют по одному принципу, хотя могут отличаться в деталях. Основой для разработки технологии редактирования генома послужила простая и относительная компактная система с единственным большим белком Cas9, который в одиночку справляется со всеми задачами. Только в случае геномного редактирования активный комплекс включает участок, соответствующий уже не вирусной ДНК, а целевому гену (например, с мутацией). Разрыв, который образуется в клеточной ДНК, можно использовать для исправления нежелательной мутации либо встраивания трансгена.
Системы CRISPR-Cas, позволившие манипулировать хромосомной ДНК непосредственно в живых клетках, стали основой революционной биотехнологии, с помощью которой можно не только создавать новые породы и сорта сельскохозяйственных животных и растений, но и, в недалеком будущем, лечить наследственные заболевания человека. Так изучение стратегий борьбы бактерий с бактериофагами, несмотря на свою кажущуюся фундаментальность, принесло неоценимую пользу человечеству.
Для борьбы с вредителями и болезнями растений используют грибы, бактерии, вирусы, гельминтов и простейших.
1. Патогенность, способность вызывать болезненные явления.
2. Вирулентность – быстро размножаться в тканях насекомого и вызывать смерть.
3. Способность проникать в организм насекомого пассивно (с пищей) и активно (через покровы тела).
4. Сохранность в природе.
Использование грибов.Наиболее патогенны для насекомых фикомицеты – облигатные паразиты. Известно около 400 видов энтомопатогенных грибов. Многие лесные насекомые поражаются плесенью гриба белая мюскардина (препарат боверин). Кроме того, различают розовую и зеленую мюскардину (в зависимости от цвета мицелия). Эти грибы вызывают мускардиноз насекомых (погибшие насекомые уменьшаются в размерах, сморщиваются и покрываются белым мучнистым налетом, состоящим из грибницы и конидий). Для массового выращивания грибов применяют твердые среды растительного происхождения (картофель, кукурузу, мельничные отходы), которые стерилизуют под давлением и затем засевают соответствующей грибной культурой. Грибы выращивают при определенной температуре, высушивают, размалывают и используют в виде препаратов. Микозы (грибные заболевания) не являются острозаразными, в естественных условиях не вызывают эпизоотий и поражают, в основном, ослабленных насекомых.
Использование бактерий получило широкое распространение. Они применяются в виде бактериальных препаратов. Технология их приготовления заключается в массовом накоплении спор путем стерильного выращивания бактерий в жидких средах. Затем споры отделяют от жидкости сепарированием, смешивают с нейтральным наполнителем и высушивают. Бактериальные препараты не имеют специфического запаха, не отпугивают насекомых, не повреждают растений, практически неопасны для людей и животных. Они готовятся на основе спорообразующих кристаллоносных энтомопатогенных бацилл группы бациллюс турингиензис (Bacillus thuringiensis Berliner). Характерным для этой группы бацилл является образование в спорангиях рядом со спорой протеиновых кристаллов белкового токсина очень токсичных для насекомых, в основном для чешуекрылых.
Наряду с эндотоксином бактерии в процессе жизнедеятельности вырабатывают экзотоксин, по химической природе близкий к нуклеотидам. Он накапливается в культуральной жидкости после отделения от нее спор и кристаллов бактерии и вызывает специфические задержки линьки у личинок насекомых и уродства у имаго, токсичен для многих групп насекомых.
Поражение гусениц вредителей происходит при попадании спор и кристаллов в кишечник вместе с пищей. Проглотив первые порции корма, гусеницы прекращают питание вследствие действия эндотоксина. Затем начинается размножение попавших внутрь тела гусениц бацилл, сопровождающееся образованием токсичных для насекомых веществ (главным образом экзотоксинов).
У погибших гусениц наблюдается разложение внутреннего содержимого: сквозь легко разрывающиеся покровы вытекает бурая жидкость, содержащая споры бацилл и кристаллы эндотоксина.
Бактериальные препараты выпускаются промышленностью в виде смачивающихся порошков и паст. В 1 г порошка содержится не менее 30 млрд. спор и столько же кристаллов, в 1 г пасты – не менее 20 млрд. спор. Гарантийный срок хранения 1 год. Инертным наполнителем обычно служит каолин. Бактериальные препараты применяют методом мелкокапельного авиационного или наземного опрыскивания насаждений.
В зависимости от температурных условий (при температуре ниже 18°С развитие бацилл сильно замедляется, отмирание гусениц затягивается и многие из них выздоравливают), активности препаратов и физиологического состояния вредителя гибель гусениц начинается по истечении 2-3 суток после обработки, а массовая смертность обычно бывает в интервале от 3-5 до 7-10 дней.
Эффективность препаратов во многом зависит от дисперсности дробления рабочей жидкости, определяющей густоту покрытия обрабатываемой поверхности растений.
Широкое применение имеют: лепидоцид (против соснового шелкопряда, непарного шелкопряда, златогузки, шелкопряда-монашенки), лепидобактоцид (хвоелистогрызущих гусениц младших возрастов), битоксибациллин против листоверток, шелкопряда, пядениц, златогузки). Лепидоцид, препарат, вырабатываемый на основе бактерии Bacillus thuringiensis, var. kurstaki (спорово-кристаллический комплекс). Битоксибациллин - экзотоксин Bacillus thuringiensis, var. thuringiensis(спорово-кристаллический комплекс).

Эффективность бактериальных препаратов при различных способах использования в борьбе с вредителями колеблется в пределах 70—90%.
Использование вирусов. Вирусы вызывают заболевания (вирозы) на стадии личинки у многих видов чешуекрылых и пилильщиков. Возникающие очаги инфекции обычно действуют несколько лет, обеспечивая вторичное инфекцирование и гибель гусениц в течение нескольких последующих поколений.
Основная вирусная болезнь насекомых – полиэдроз (известно около 170 видов). Он сопровождается появлением в клетках различных тканей многочисленных внутриядерных белковых включений, имеющих форму многогранников (полиэдров). Полиэдры устойчивы к действию неблагоприятных факторов (химикатов, солнечного света, ферментов и др.) и сохраняют жизнеспособность находящихся в них вирусов многие годы. Размеры полиэдров колеблются в пределах 0,3-15 мкм. В белковом матриксе полиэдров расположены вирионы в форме палочек длиной 200-400 и шириной 20-50 мкм. Вирус передается через яйца (трансовариально) и специфичен для каждого вида насекомого. Больные насекомые становятся малоподвижными, прекращают питание, их покровы приобретают светлую окраску, утончаются, и через их разрывы вытекает мутная, непахнущая, беловатая жидкость.
Для приготовления вирусных препаратов зараженных полиэдрозом гусениц высушивают, затем их содержимое соответствующим образом обрабатывают. Вирусные препараты используют для борьбы с сибирским шелкопрядом (вирин-ГСШ), сосновыми пилильщиками (вирин-Диприон), шелкопрядом монашенкой (вирин-ПШМ).
Вирин-Диприон представляет собой суспензию, состоящую из полиэдров. Ее разбавляют водой и используют для опрыскивания очагов сосновых пилильщиков.

Обзор
Автор
Редакторы
Обратите внимание!
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science.
Эволюция и происхождение вирусов
В 2007 году сотрудники биологического факультета МГУ Л. Нефедова и А. Ким описали, как мог появиться один из видов вирусов — ретровирусы. Они провели сравнительный анализ геномов дрозофилы D. melanogaster и ее эндосимбионта (микроорганизма, живущего внутри дрозофилы) — бактерии Wolbachia pipientis. Полученные данные показали, что эндогенные ретровирусы группы gypsy могли произойти от мобильных элементов генома — ретротранспозонов. Причиной этому стало появление у ретротранспозонов одного нового гена — env, — который и превратил их в вирусы. Этот ген позволяет вирусам передаваться горизонтально, от клетки к клетке и от носителя к носителю, чего ретротранспозоны делать не могли. Именно так, как показал анализ, ретровирус gypsy передался из генома дрозофилы ее симбионту — вольбахии [7]. Это открытие упомянуто здесь не случайно. Оно нам понадобится для того, чтобы понять, чем вызваны трудности борьбы с вирусами.
Из давних письменных источников, оставленных историком Фукидидом и знахарем Галеном, нам известно о первых вирусных эпидемиях, возникших в Древней Греции в 430 году до н.э. и в Риме в 166 году. Часть вирусологов предполагает, что в Риме могла произойти первая зафиксированная в источниках эпидемия оспы. Тогда от неизвестного смертоносного вируса по всей Римской империи погибло несколько миллионов человек [8]. И с того времени европейский континент уже регулярно подвергался опустошающим нашествиям всевозможных эпидемий — в первую очередь, чумы, холеры и натуральной оспы. Эпидемии внезапно приходили одна за другой вместе с перемещавшимися на дальние расстояния людьми и опустошали целые города. И так же внезапно прекращались, ничем не проявляя себя сотни лет.
Вирус натуральной оспы стал первым инфекционным носителем, который представлял действительную угрозу для человечества и от которого погибало большое количество людей. Свирепствовавшая в средние века оспа буквально выкашивала целые города, оставляя после себя огромные кладбища погибших. В 2007 году в журнале Национальной академии наук США (PNAS) вышла работа группы американских ученых — И. Дэймона и его коллег, — которым на основе геномного анализа удалось установить предположительное время возникновения вируса натуральной оспы: более 16 тысяч лет назад. Интересно, что в этой же статье ученые недоумевают по поводу своего открытия: как так случилось, что, несмотря на древний возраст вируса, эпидемии оспы не упоминаются в Библии, а также в книгах древних римлян и греков [9]?
Строение вирусов и иммунный ответ организма

Рисунок 1. Первооткрыватель вирусов Д.И. Ивановский (1864–1920) (слева) и английский врач Эдвард Дженнер (справа).

Почти все известные науке вирусы имеют свою специфическую мишень в живом организме — определенный рецептор на поверхности клетки, к которому и прикрепляется вирус. Этот вирусный механизм и предопределяет, какие именно клетки пострадают от инфекции. К примеру, вирус полиомиелита может прикрепляться лишь к нейронам и потому поражает именно их, в то время как вирусы гепатита поражают только клетки печени. Некоторые вирусы — например, вирус гриппа А-типа и риновирус — прикрепляются к рецепторам гликофорин А и ICAM-1, которые характерны для нескольких видов клеток. Вирус иммунодефицита избирает в качестве мишеней целый ряд клеток: в первую очередь, клетки иммунной системы (Т-хелперы, макрофаги), а также эозинофилы, тимоциты, дендритные клетки, астроциты и другие, несущие на своей мембране специфический рецептор СD-4 и CXCR4-корецептор [13–15].

Одновременно с этим в организме реализуется еще один, молекулярный, защитный механизм: пораженные вирусом клетки начинают производить специальные белки — интерфероны, — о которых многие слышали в связи с гриппозной инфекцией. Существует три основных вида интерферонов. Синтез интерферона-альфа (ИФ-α) стимулируют лейкоциты. Он участвует в борьбе с вирусами и обладает противоопухолевым действием. Интерферон-бета (ИФ-β) производят клетки соединительной ткани, фибробласты. Он обладает таким же действием, как и ИФ-α, только с уклоном в противоопухолевый эффект. Интерферон-гамма (ИФ-γ) синтезируют Т-клетки (Т-хелперы и (СD8+) Т-лимфоциты), что придает ему свойства иммуномодулятора, усиливающего или ослабляющего иммунитет. Как именно интерфероны борются с вирусами? Они могут, в частности, блокировать работу чужеродных нуклеиновых кислот, не давая вирусу возможности реплицироваться (размножаться).

Причины поражений в борьбе с ВИЧ
Тем не менее нельзя сказать, что ничего не делается в борьбе с ВИЧ и нет никаких подвижек в этом вопросе. Сегодня уже определены перспективные направления в исследованиях, главные из которых: использование антисмысловых молекул (антисмысловых РНК), РНК-интерференция, аптамерная и химерная технологии [12]. Но пока эти антивирусные методы — дело научных институтов, а не широкой клинической практики*. И потому более миллиона человек, по официальным данным ВОЗ, погибают ежегодно от причин, связанных с ВИЧ и СПИДом.

Подобный вирусный механизм характерен не только для ВИЧ. Он описан и при инфицировании некоторыми другими опасными вирусами: такими, как вирусы Денге и Эбола. Но при ВИЧ антителозависимое усиление инфекции сопровождается еще несколькими факторами, делая его опасным и почти неуязвимым. Так, в 1991 году американские клеточные биологи из Мэриленда (Дж. Гудсмит с коллегами), изучая иммунный ответ на ВИЧ-вакцину, обнаружили так называемый феномен антигенного импринтинга [23]. Он был описан еще в далеком 1953 году при изучении вируса гриппа. Оказалось, что иммунная система запоминает самый первый вариант вируса ВИЧ и вырабатывает к нему специфические антитела. Когда вирус видоизменяется в результате точечных мутаций, а это происходит часто и быстро, иммунная система почему-то не реагирует на эти изменения, продолжая производить антитела к самому первому варианту вируса. Именно этот феномен, как считает ряд ученых, стоит препятствием перед созданием эффективной вакцины против ВИЧ.

Открытие биологов из МГУ — Нефёдовой и Кима, — о котором упоминалось в самом начале, также говорит в пользу этой, эволюционной, версии.

Сегодня не только ВИЧ представляет опасность для человечества, хотя он, конечно, самый главный наш вирусный враг. Так сложилось, что СМИ уделяют внимание, в основном, молниеносным инфекциям, вроде атипичной пневмонии или МЕRS, которыми быстро заражается сравнительно большое количество людей (и немало гибнет). Из-за этого в тени остаются медленно текущие инфекции, которые сегодня гораздо опаснее и коварнее коронавирусов* и даже вируса Эбола. К примеру, мало кто знает о мировой эпидемии гепатита С, вирус которого был открыт в 1989 году**. А ведь по всему миру сейчас насчитывается 150 млн человек — носителей вируса гепатита С! И, по данным ВОЗ, каждый год от этой инфекции умирает 350-500 тысяч человек [33]. Для сравнения — от лихорадки Эбола в 2014-2015 гг. (на состояние по июнь 2015 г.) погибли 11 184 человека [34].
* — Коронавирусы — РНК-содержащие вирусы, поверхность которых покрыта булавовидными отростками, придающими им форму короны. Коронавирусы поражают альвеолярный эпителий (выстилку легочных альвеол), повышая проницаемость клеток, что приводит к нарушению водно-электролитного баланса и развитию пневмонии.

Рисунок 8. Электронная микрофотография воссозданного вируса H1N1, вызвавшего эпидемию в 1918 г. Рисунок с сайта phil.cdc.gov.
Почему же вдруг сложилась такая ситуация, что буквально каждый год появляются новые, всё более опасные формы вирусов? По мнению ученых, главные причины — это сомкнутость популяции, когда происходит тесный контакт людей при их большом количестве, и снижение иммунитета вследствие загрязнения среды обитания и стрессов. Научный и технический прогресс создал такие возможности и средства передвижения, что носитель опасной инфекции уже через несколько суток может добраться с одного континента на другой, преодолев тысячи километров.
Вирусы уже очень давно изучают в лабораториях, а также выводят полученные с нуля виды или иначе говоря de novo. Однако работа вирусологов массово заинтересовала широкую общественность только после начала пандемии коронавируса. Рассказываем, с какой целью выводят биологическое оружие, есть ли у модифицированных вирусов положительные задачи и что известно о происхождении коронавируса.
Искусственные вирусы
Многие вирусы могут быть получены de novo, то есть с нуля, а первый искусственный вирус был создан в 2002 году. Несмотря на некоторые неправильные трактовки, при этом процессе синтезируется не сам вирус как таковой, а его геномная ДНК (в случае ДНК-вирусов) или комплементарная копия ДНК его генома (в случае РНК-вирусов).
У вирусов многих семейств искусственная ДНК или РНК, если она введена в клетку, проявляет инфекционные свойства. Иными словами, такие вирусы содержат всю необходимую информацию для образования новых вирусов.
Эту технологию в настоящее время используют для разработки вакцин нового типа. Возможность создавать искусственные вирусы имеет далеко идущие последствия, поскольку вирус не может вымереть, пока известна его геномная последовательность и имеются чувствительные к нему клетки.
В наши дни полные геномные последовательности 2408 различных вирусов (в том числе оспы) находятся в публичном доступе в онлайн-базе данных, поддерживаемой Национальными институтами здравоохранения США.

Микрофотография, показывающая цитопатические эффекты, вызванные вирусом простого герпеса первого типа. Тест Папаниколау
Как из вирусов делают биологическое оружие
Способность вирусов вызывать опустошительные эпидемии среди людей порождает беспокойство, что вирусы могут использоваться как биологическое оружие.
Биологическое оружие — это патогенные микроорганизмы или их споры, вирусы, бактериальные токсины, заражающие людей и животных, предназначенные для массового поражения живой силы и населения противника, сельскохозяйственных животных, посевов сельскохозяйственных культур, заражения продовольствия и источников воды, а также порчи некоторых видов военного снаряжения и военных материалов.
Биологическое оружие включает также средства доставки патогенных микроорганизмов и животных-переносчиков. Является оружием массового поражения и запрещено согласно Женевскому протоколу 1925 года.
Дополнительные опасения вызвало успешное воссоздание вредоносного вируса испанского гриппа в лаборатории. Другим примером может служить вирус оспы. Он на всем протяжении истории опустошал множество стран вплоть до его окончательного искоренения. Официально образцы вируса оспы хранятся лишь в двух местах в мире — в двух лабораториях в России и США.
Опасения, что он может быть использован как оружие, не совсем беспочвенны; вакцина против оспы иногда имеет тяжелые побочные эффекты — в последние годы до официально объявленного искоренения вируса больше людей серьёзно заболели из-за вакцины, чем от вируса, поэтому вакцинация против оспы больше не практикуется повсеместно. По этой причине большая часть современного населения Земли практически не имеет устойчивости к оспе.
Способы применения бактериальных и вирусных средств
Средствами доставки и способами применения биологического оружия, как правило, являются:
- боевые части ракет;
- авиационные бомбы;
- артиллерийские мины и снаряды;
- пакеты (мешки, коробки, контейнеры), сбрасываемые с самолетов;
- специальные аппараты, рассеивающие насекомых с самолетов;
- диверсионные методы.
Заболевание в этом случае может произойти в результате прямого контакта с заражёнными предметами. Возможно также преднамеренное оставление при отходе инфекционных больных с тем, чтобы они явились источником заражения.
При разрыве боеприпасов, снаряженных бактериальной рецептурой, образуется бактериальное облако, состоящее из взвешенных в воздухе мельчайших капелек жидкости или твёрдых частиц. Облако, распространяясь по ветру, рассеивается и оседает на землю, образуя заражённый участок, площадь которого зависит от количества рецептуры, её свойств и скорости ветра.
Проблемой является то, что вне природного очага обитания и без соответствующих его экологической обстановке механизмов передачи, возбудитель заболевания передаваться людям не будет.
Особенности поражения биологическим оружием
При поражении бактериальными или вирусными средствами заболевание наступает не сразу, почти всегда имеется скрытый (инкубационный) период, в течение которого заболевание не проявляет себя внешними признаками, а пораженный не теряет боеспособности.
Некоторые заболевания (чума, холера, сибирская язва) способны передаваться от больного человека здоровому и, быстро распространяясь, вызывать эпидемии. Установить факт применения бактериальных средств и определить вид возбудителя достаточно трудно, поскольку ни микробы, ни токсины не имеют ни цвета, ни запаха, ни вкуса, а эффект их действия может проявиться через большой промежуток времени.
Обнаружение бактерий и вирусов возможно только путём проведения специальных лабораторных исследований, на что требуется значительное время, что затрудняет своевременное проведение мероприятий по предупреждению эпидемических заболеваний.
Признаком применения бактериологического оружия являются также валяющиеся на местности использованные боеприпасы (предназначенные для него) и другие средства его доставки. В некоторых случаях также — внезапное появление или резкое увеличение количества определенных насекомых или грызунов (например — блохи на снегу).
Современные стратегические средства биологического оружия используют смеси вирусов и спор бактерий для увеличения вероятности летальных исходов при применении, однако используются, как правило, штаммы, не передающиеся от человека к человеку, чтобы территориально локализовать их воздействие и избежать вследствие этого собственных потерь.
Искусственные вирусы можно использовать во благо
Ученые из NPL (Национальной физической лабораторией), работая с партнерами из Кембриджского и Эксетерского университетов, а также из Королевского колледжа Лондона, разработали создают искусственный вирус для борьбы с супербактериями
Рост числа супербактерий вызывает серьезную озабоченность в медицинском сообществе, поскольку бактерии эволюционируют, чтобы избежать существующих методов лечения быстрее, чем разрабатывают новые антибиотики. Вместо того чтобы создавать новые лекарства, группа экспертов пошла другим путем.
Авторы работы, опираясь на принципы архитектуры вируса, создали синтетический белок Ψ—капсид, который собирается из небольшого молекулярного мотива или характерной последовательности нуклеотидов (в ДНК, РНК) или аминокислот (в белках). Мотив нашли в клетках человека. Он может распознавать молекулярные паттерны, связанные с патогенами, на бактериальных поверхностях.
Отмечается, что благодаря комбинации наноразмерных и одноклеточных изображений, команда подтвердила, что капсиды наносят непоправимый ущерб бактериям. Капсиды были одинаково эффективны в любой из своих хиральных форм, что может сделать их невидимыми для иммунной системы хозяина, а также позволит убивать различные фенотипы бактерий и супербактерий без цитотоксичности in vitro и in vivo.
Коронавирус создан искусственно?
Пока нельзя сказать однозначно. Предлагаем ознакомиться с недавним необычном исследованием, в котором британский и норвежский ученые Ангус Далглиш и Биргер Сёренсен утверждают, что COVID-19 был создан в лаборатории.
По их словам, ранее они уже пытались опубликовать результаты своего исследования, но были отвергнуты научными журналами, которые были уверены в естественном происхождении вируса. Сейчас в ряде стран снова заговорили о необходимости пересмотреть версии возникновения COVID-19, указывает издание.
Исследователи обратили внимание на эксперименты, проведенные в лаборатории в Ухане в период с 2002 по 2019 год, и выяснили, что
их китайские коллеги, некоторые из которых работают совместно с американскими университетами, занимались исследованиями по изменению вируса таким образом, чтобы повысить его заразность.
Бактериофаги – это вирусы, которые поражают только бактерий. В ходе инфекции они влияют на все процессы жизнедеятельности бактериальной клетки, фактически превращая ее в фабрику по производству вирусного потомства. В конце концов клетка разрушается, а вновь образованные вирусные частицы выходят наружу и могут заражать новые бактерии.
Несмотря на огромное число и разнообразие природных фагов, встречаемся мы с ними редко. Однако бывают ситуации, когда деятельность этих вирусов не остается незамеченной. Например, на предприятиях, где производят сыры, йогурты и другие молочно-кислые продукты, часто приходится сталкиваться с вирусной атакой на бактерии, сбраживающие молоко. В большинстве таких случаев фаговая инфекция распространяется молниеносно, и полезные бактерии гибнут, что приводит к значительным экономическим потерям (Neve et al., 1994).
Именно благодаря прикладным исследованиям в интересах молочной промышленности, направленным на получение устойчивых к бактериофагам штаммов молочно-кислых бактерий, был открыт ряд механизмов, с помощью которых бактерии избегают инфекции. Параллельно были изучены способы, с помощью которых вирусы, в свою очередь, преодолевают бактериальные системы защиты (Moineau et al., 1993).
Кто защищен – тот вооружен
На сегодня известно пять основных, весьма хитроумных механизмов защиты, которые бактерии выработали в непрестанной борьбе с вирусами: изменение рецептора на поверхности клетки; исключение суперинфекции; системы абортивной инфекции; системы рестрикции-модификации и, наконец, системы CRISPR-Cas.
К средствам противовирусной защиты бактерий относятся и системы рестрикции-модификации, в которые входят гены, кодирующие два белка-фермента – рестриктазу и метилазу. Рестриктаза узнает определенные последовательности ДНК длиной 4—6 нуклеотидов и вносит в них двуцепочечные разрывы. Метилаза, напротив, ковалентно модифицирует эти последовательности, добавляя к отдельным нуклеотидным основаниям метильные группы, что предотвращает их узнавание рестриктазой.
Врага нужно знать в лицо
Системы CRISPR-Cas являются уникальным примером адаптивного иммунитета бактерий. При проникновении в клетку ДНК фага специальные белки Cas встраивают фрагменты вирусной ДНК длиной 25—40 нуклеотидов в определенный участок генома бактерии (Barrangou et al., 2007). Такие фрагменты называются спейсерами (от англ. spacer – промежуток), участок, где происходит встраивание, – CRISPR-кассета (от англ. Clustered Regularly Interspaced Short Palindromic Repeats), а сам процесс приобретения спейсеров – адаптацией.
Чтобы использовать спейсеры в борьбе с фаговой инфекцией, в клетке должен происходить еще один процесс, управляемый белками Cas, названный интерференцией. Суть его в том, что в ходе транскрипции CRISPR-кассеты образуется длинная молекула РНК, которая разрезается белками Cas на короткие фрагменты – защитные криспрРНК (крРНК), каждая из которых содержит один спейсер. Белки Cas вместе с молекулой крРНК образуют эффекторный комплекс, который сканирует всю ДНК клетки на наличие последовательностей, идентичных спейсеру (протоспейсеров). Найденные протоспейсеры расщепляются белками Cas (Westra et al., 2012; Jinek et al., 2012).
Системы CRISPR-Cas обнаружены у большинства прокариот – бактерий и архей. Хотя общий принцип действия всех известных систем CRISPR-Cas одинаков, механизмы их работы могут существенно отличаться в деталях. Наибольшие различия проявляются в строении и функционировании эффекторного комплекса, в связи с чем системы CRISPR-Cas делят на несколько типов. На сегодняшний день описаны шесть типов таких неродственных друг другу систем (Makarova et al., 2015; Shmakov et al., 2015).
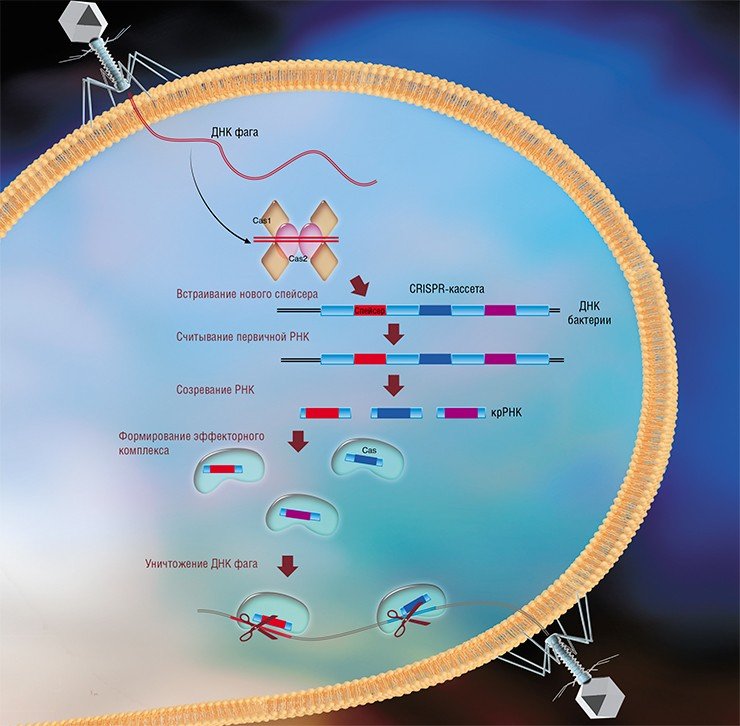
Наиболее изученной является система CRISPR-Cas I типа, которой обладает излюбленный объект молекулярно-биологических исследований – бактерия кишечная палочка (Esсherichia coli). Эффекторный комплекс в этой системе состоит из нескольких небольших белков Cas, каждый из которых отвечает за разные функции: разрезание длинной некодирующей CRISPR РНК, связывание коротких крРНК, поиск, а затем разрезание ДНК-мишени.
Гонка вооружений
Бактериофаги, как факторы среды, вызывают направленные изменения в геноме бактерий, которые наследуются и дают бактериям явное преимущество, спасая от повторных инфекций. Поэтому системы CRISPR-Cas можно считать примером ламарковской эволюции, при которой происходит наследование благоприобретенных признаков (Koonin et al., 2009)
Некоторые бактериофаги реагируют на наличие в бактериальной клетке систем CRISPR-Cas выработкой особых анти CRISPR-белков, способных связываться с белками Cas и блокировать их функции (Bondy-Denomy et al., 2015). Еще одно ухищрение — обмен участков генома вируса, на которые нацелена система CRISPR-Cas, на участки геномов родственных вирусов, отличающихся по составу нуклеотидной последовательности (Paez-Espino et al., 2015).
Благодаря постоянному совершенствованию биоинформатических алгоритмов поиска, а также включению в анализ все большего количества прокариотических геномов, открытие новых типов CRISPR-Cas систем является делом недалекого будущего. Предстоит также выяснить и детальные механизмы работы многих недавно открытых систем. Так, в статье, опубликованной в 2016 г. в журнале Science и посвященной анализу системы CRISPR-Cas VI типа, описан белок С2с2, образующий эффекторный комплекс с крРНК, который нацелен на деградацию не ДНК, а РНК (Abudayyeh et al., 2016). В будущем такое необычное свойство может быть использовано в медицине для регулирования активности генов путем изменения количества кодируемых ими РНК.
Изучение стратегий борьбы бактерий с бактериофагами, несмотря на свою кажущуюся фундаментальность и отвлеченность от задач практической медицины, принесло неоценимую пользу человечеству. Примерами этого могут служить методы молекулярного клонирования и редактирования геномов – направленного внесения или удаления мутаций и изменения уровня транскрипции определенных генов.
Благодаря быстрому развитию методов молекулярной биологии всего лишь через несколько лет после открытия механизма действия систем CRISPR-Cas была создана работающая технология геномного редактирования, способная бороться с болезнями, ранее считавшимися неизлечимыми. Доступность и простота этой технологии позволяют рассматривать ее как основу для медицины, ветеринарии, сельского хозяйства и биотехнологий будущего, которые будут базироваться на направленных и безопасных генных модификациях.
Нет никаких сомнений, что дальнейшее изучение взаимодействия бактерий и их вирусов может открыть перед нами такие возможности, о которых мы сейчас даже не подозреваем.
Abudayyeh O. O., Gootenberg J. S., Konermann S. et al. C 2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector // Science. 2016. V. 353: aaf5573.
Barrangou R., Fremaux C., Deveau H. et al. CRISPR provides acquired resistance against viruses in prokaryotes // Science. 2007. V. 315. P. 1709–1712.
Bikard D., Marraffini L. A. Innate and adaptive immunity in bacteria: mechanisms of programmed genetic variation to fight bacteriophages // Curr. Opin. Immunol. 2012. V. 1 P. 15–20.
Bondy-Denomy J., Garcia B., Strum S. et al. Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins // Nature. 2015. V. 526. P. 136–139.
Calendar R., Abedon S. T. The Bacteriophages // 2nd Ed., Oxford University Press. 2006.
Datsenko K. A., Pougach K., Tikhonov A. et al. Molecular memory of prior infections activates the CRISPR/Cas adaptive bacterial immunity system // Nat. Commun. 2012. V. 3. P. 945
Jiang W., Marraffini L. A. CRISPR-Cas: New Tools for Genetic Manipulations from Bacterial Immunity Systems // Annu. Rev. Microbiol. 2015. V. 69. P. 209–28.
Jinek M., Chylinski K., Fonfara I., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity // Science. 2012. V. 337. P. 816–821.
Koonin E. V., Wolf Y. I. Is evolution Darwinian or/and Lamarckian? // Biol. Direct. 2009. V. 4. P. 42.
Lopez-Pascua L., Buckling A. Increasing productivity accelerates host-parasite coevolution // J. Evol. Biol. 2008. V. 3. P. 853–860.
Makarova K. S., Wolf Y. I., et al. An updated evolutionary classification of CRISPR-Cas systems // Nat. Rev. Microbiol. 2015. V. 11. P. 722–736.
Moineau, S., Pandian S., Klaenhammer T. R. Restriction/modification systems and restriction endonucleases are more effective on lactococcal bacteriophages that have emerged recently in the dairy industry // Appl. Envir. Microbiol. 1993. V. 59. P. 197–202.
Neve H., Kemper U., et al. Monitoring and characterization of lactococcal bacteriophage in a dairy plant // Kiel. Milckwirtsch. Forschungsber. 1994. V. 46. P. 167–178.
Nuñez J. K., Harrington L. B., et al. Foreign DNA capture during CRISPR-Cas adaptive immunity // Nature. 2015a. V. 527. P. 535–538.
Nuñez J. K., Kranzusch P. J., et al. Cas1-Cas2 complex formation mediates spacer acquisition during CRISPR-Cas adaptive immunity // Nat. Struct. Mol. Biol. 2014. V. 21. P. 528–534.
Nuñez J. K., Lee A. S., Engelman A., Doudna J. A. Integrase-mediated spacer acquisition during CRISPR-Cas adaptive immunity // Nature. 2015b. V. 519. P. 193–198.
Paez-Espino D., Sharon I., et al. CRISPR Immunity Drives Rapid Phage Genome Evolution in Streptococcus thermophilus // MBio. 2015. V. 6: e00262–15.
Shmakov S., Abudayyeh O. O., Makarova K. S., et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. // Mol. Cell. 2015. V. 60. P. 385–397
Tan D., Svenningsen S. L., Middelboe M. Quorum sensing determines the choice of antiphage defense strategy in Vibrio anguillarum. // mBio 2015. V. 6: e00627–15.
Westra E. R., van Erp P. B., Künne T., et al. CRISPR immunity relies on the consecutive binding and degradation of negatively supercoiled invader DNA by Cascade and Cas3 // Mol. Cell. 2012. V. 46. P. 595–605.
Читайте также: