
Вирусы с липопротеидной оболочкой формируются
Обновлено: 19.04.2024
ГЕРПЕСА ВИРУСЫ — род вирусов, вирионы которых состоят из нуклеокапсида с икосаэдральным типом симметрии и липопротеидной оболочки. Диаметр вириона 100— 150 нм, геном представлен двухнитчатой ДНК.
Большинство известных вирусов герпеса, выделяемых от человека и животных (табл. 1), вызывает инфекционные заболевания, другие индуцируют развитие опухолей или ассоциируют с ними (табл. 2).
Вирусы простого герпеса (ВПГ — Herpes simplex virus — HSV) — одни из распространенных вирусов, поражающих человека. Современная отечественная классификация ВПГ разработана А. К. Шубладзе с сотр. на основе антигенной структуры вирусов и включает 6 групп. Все вирусы патогенны для лабораторных животных, куриных эмбрионов; в инфицированных клетках формируются внутриклеточные включения.В клеточных культурах Г. в. проявляют гемадсорбирующий эффект, образуют симп ласты; под агаровым покрытием формируют различные по морфологии бляшки. Цитопатический эффект проявляют в основном штаммы первой антигенной группы.
Позднее на основании серол, тестов гибридизации нуклеиновой к-ты и различий в нуклеотидном составе, а также по биол, свойствам ВПГ разделены на две группы: тип 1 и тип 2.

Рис. 1 Электронограмма зрелого вириона вируса простого герпеса: 1 — внешняя оболочка; 2 — центрально расположенный нуклеокапсид. Негативное контрастирование; X 150 000.

Рис. 2. Электронограмма двух незрелых: вирионов вируса простого герпеса без. наружной оболочки (указаны стрелками). Негативное контрастирование; X 200 000.

Рис. 3. Электронограмма адсорбции вириона (1) вируса простого герпеса на клетке (2). Ультратонкий срез клетки; X 100 000.

Рис. 4. Электронограмма формирования вируса простого герпеса в ядре инфицированной клетки: 1 — нуклеокапсиды; 2 — пустые (без нуклеоида) капсиды. Ультратонкий срез клетки; х 20 000.
Адсорбция вирусов на клетки тканевых культур (рис. 3) происходит в течение 2—3 час. Механизм процесса объясняется наличием ионизированных групп на поверхности клетки и вируса как неспецифической реакцией, так и специфическим взаимодействием рецепторов клетки с рецепторами вируса.

Рис. 5. Электронограмма незрелых вирионов: 1 — нуклеокапсиды (без оболочки) вируса простого герпеса в ядре клетки; 2 — зрелые вирионы — нуклеокапсиды. покрытые оболочкой, в перинуклеарном пространстве. Ультратонкий срез клетки; X 50 000.

Рис. 6. Электронограмма выхода зрелого вируса (1) из клетки (2). Ультратонкий срез клетки; X 150 000.
Сборка нуклеокапсидов (незрелых вирионов) происходит в ядре клеток (рис. 4). Формирование вируса заканчивается в процессе выхода из ядра в цитоплазму путем приобретения наружной оболочки за счет части ядерной мембраны (рис. 5). После этого зрелый вирус выходит за пределы клетки (рис. 6).
В процессе синтеза вируса происходят значительные изменения митотического режима клеток и появление колхициноподобных метафаз. Хромосомные повреждения выражаются в основном в хроматидных и изохроматидных разрывах. В ядрах происходит перегруппировка (маргинация) хроматина, набухание ядрышек с последующим лизисом и формирование эозинофильных ДНК-содержащих включений. Клетки округляются, среди них появляются сим-пласты. Далее происходит вакуолизация клеток, нарушение структуры цитоплазматических органоидов и в конечном итоге (через 24—72 часа после заражения) наступает разрушение зараженных клеток.
Г. в. являются слабым индуктором интерферона, высокочувствительны к эфиру, трипсину, прогреванию, устойчивы к рентгеновским лучам, хорошо сохраняются при низких температурах и в высушенном состоянии.
При первичном инфицировании (чаще в раннем детстве) Г. в. попадают в кровь (вирусемия), размножаются в тканях экто- и эндодермального происхождения и вызывают различные острые клинические проявления, наиболее часто связанные с поражением слизистых оболочек, кожи, роговицы, иногда печени, мозга и других органов. В большей части случаев наступает клиническое выздоровление, однако вирус (или его геном) сохраняется в организме в течение всей жизни в скрытой форме при отсутствии каких-либо симптомов заболевания (ок. 90% населения являются носителями ВПГ). Под влиянием некоторых стрессовых факторов (переохлаждение, различные бактериальные инфекции и т. д.) вирус периодически реактивируется, вызывая патол. процесс, напоминающий первичное проявление инфекции. В результате вирусемии, связанной с наличием вируса во всех фракциях крови, в момент заболевания (возможно и в межрецидивном периоде) вирус может преодолевать плацентарный барьер, проникать в плод и вызывать различные патол, процессы. Особенностью хрон, герпетической инфекции является постоянное наличие в крови человека специфических вируснейтрализующих антител.
В экспериментах на клеточных культурах показано, что при хрон, герпетической инфекции синтез вируса происходит по типу продуктивной инфекции в отдельных клетках.
Длительная персистенция Г. в. в организме создает возможность трансформирующего действия вируса на клетки. В экспериментальных условиях феномен трансформации установлен для Г. в. животных и человека. Известны Онкогенные Г. в., выделенные от лягушек, птиц, грызунов, крупного рогатого скота и некоторых видов обезьян. Имеются данные о выделении Г. в. и герпесоподобных вирусов из опухолей человека. При этом выявлена интимная ассоциация Г. в. с опухолевыми клетками, однако окончательная роль Г. в. в развитии новообразований у человека остается недоказанной.
Таблица 1. ВИРУСЫ ГЕРПЕСА, ВЫДЕЛЯЕМЫЕ ОТ ЧЕЛОВЕКА И ЖИВОТНЫХ
Вирусы простого герпеса Вирусы ветряной оспы, опоясывающего лишая Вирус цитомегалии
Структура и состав вирусов. Строение вирусов и вирусных частиц.
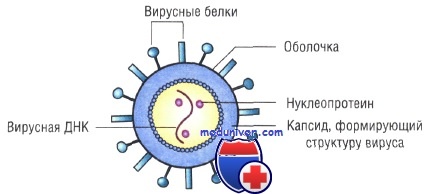
Основным структурным компонентом вирионов (полных вирусных частиц) является нуклеокапсид, т.е. белковый чехол (капсид) в котором заключен вирусный геном (ДНК или РНК). Нуклеокапсид большинства семейств вирусов окружен липопротеиновой оболочкой. Между оболочкой и нуклеокапсидом у некоторых вирусов (орто-, парамиксо-, рабдо-, фило- и ретровирусов) находится негликозилированный матриксный белок, придающий дополнительную жесткость вирионам. Вирусы большинства семейств имеют оболочку, которая играет важную роль в инфекционности. Наружный слой оболочки вирионы приобретают, когда нуклеокапсид проникает через клеточную мембрану почкованием.
Белки оболочки кодируются вирусом, а липиды заимствуются из мембраны клетки. Гликопротеины обычно в виде димеров и тримеров образуют пепломеры (выступы) на поверхности вирионов (орто-, парамиксовирусы, рабдо-, фило-, корона-, бунья-, арена-, ретровирусы). Гликозилированные белки слияния связаны с пепломерами и выполняют ключевую роль в проникновении вируса в клетку. Капсиды и оболочки вирионов образуются множеством копий одного или нескольких видов белковых субъединиц в результате процесса самосборки. Взаимодействие в системе белок-белок, благодаря слабым химическим связям, ведет к объединению симметричных капсидов.
Различия вирусов по форме и размеру вирионов зависят от формы, размера и количества структурных белковых субъединиц и природы взаимодействия между ними.
Капсид состоит из множества морфологически выраженных субъединиц (капсомеров), собранных из вирусных полипептидов строго определенным образом, в соответствии с относительно простыми геометрическими принципами. Белковые субъединицы, соединяясь друг с другом, образуют капсиды двух видов симметрии: изометрические и спиральные. Структура нуклеокапсида оболочечных вирусов сходна со структурой нуклеокапсида безоболочечных вирусов. На поверхности оболочки вирусов различают морфологически выраженные гликопротеиновые структуры — пепломеры.
В состав суперкапсидной оболочки входят липиды (до 20—35%) и углеводы (до 7—8%), имеющие клеточное происхождение. Она состоит из двойного слоя клеточных липидов и вирусспецифических белков, расположенных снаружи и изнутри липидного биослоя. Наружный слой суперкапсидной оболочки представляют пепломеры (выступы) одного или более типов, состоящие из одной или нескольких молекул гликопротеинов. Нуклеокапсид у оболочечных вирусов часто называют сердцевиной (core), а центральную часть вирионов, содержащую нуклеиновую кислоту, называют нуклеоидом.
Капсомеры (пепломеры) состоят из структурных единиц, построенных из одной либо из нескольких гомологичных или гетерологичных полипептидных цепей (белковых субъединиц).
Изометрические капсиды представляют собой не сферы, а правильные многогранники (икосаэдры). Их линейные размеры идентичны по осям симметрии. Согласно Каспару и Клугу (1962), капсомеры в капсидах расположены в соответствии с икосаэдрической симметрией.
Такие капсиды состоят из идентичных субъединиц, образующих икосаэдр. Они имеют 12 вершин (углов), 30 граней и 20 поверхностей в виде равнобедренных треугольников. В соответствии с этим правилом капсид полиовируса и вируса ящура образован 60 белковыми структурными единицами, каждая из которых состоит из четырех полипептидных цепей.
Икосаэдр оптимально решает проблему укладки повторяющихся субъединиц в строгую компактную структуру при минимальном объеме. Только некоторые конфигурации структурных субъединиц могут сформировать поверхности, образовать вершины и грани вирусного икосаэдра. Например, структурные субъединицы аденовируса на поверхностях и гранях формируют шестигранные капсомеры (гексоны), а на вершинах - пятигранные капсомеры (пептоны). У одних вирусов оба вида капсомеров образуются одними и теми же полипептидами, у других — разными полипептидами. Так как структурные субъединицы разных вирусов различаются между собой, то одни вирусы кажутся более гексагональными, другие — более сферическими.
Все известные ДНК-содержащие вирусы позвоночных, за исключением вирусов оспы, а также многие РНК-содержащие вирусы (7 семейств) имеют кубический тип симметрии капсида.
Реовирусы, в отличие от других вирусов позвоночных, имеют двойной кап-сид (наружный и внутренний), причем каждый состоит из морфологических единиц.
Вирусы, обладающие спиральным типом симметрии, имеют вид цилиндрической нитевидной структуры, их геномная РНК имеет вид спирали и находится внутри капсида. Все вирусы животных спиральной симметрии окружены липопротеиновой оболочкой.
Спиральные нуклеокапсиды характеризуются длиной, диаметром, шагом спирали и числом капсомеров, приходящихся на один оборот спирали. Так, у вируса Сендай (парамиксовирус) нуклеокапсид представляет собой спираль длиной около 1 мкм, диаметром 20 нм и шагом спирали 5 нм. Капсид состоит примерно из 2400 структурных единиц, каждая из которых является белком с молекулярной массой 60 кД. На каждый виток спирали приходится 11—13 субъединиц.
У вирусов со спиральным типом симметрии нуклеокапсида укладка белковых молекул в спираль обеспечивает максимальное взаимодействие между нуклеиновой кислотой и белковыми субъединицами. У икосаэдрических вирусов нуклеиновая кислота находится внутри вирионов в скрученном состоянии и взаимодействует с одним или несколькими полипептидами, расположенными внутри капсида.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Строение вирусов. Классификация вирусов
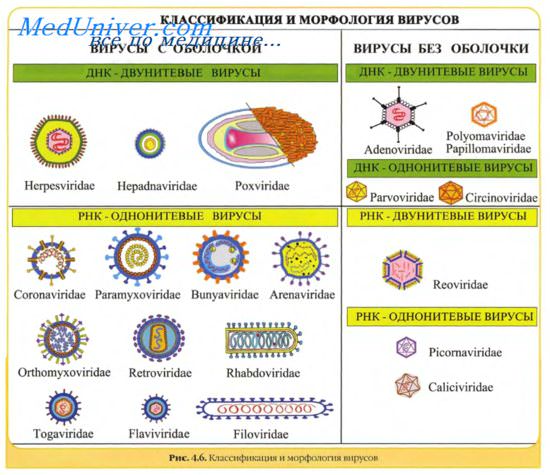
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
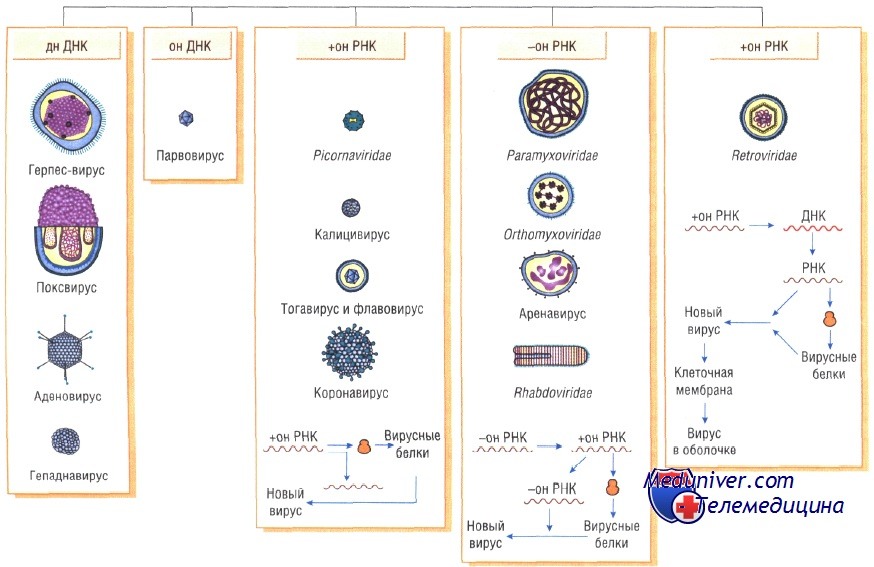
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
РЕОВИРУСЫ (Reoviridae) - семейство РНК-содержащнх вирусов, объединяющее вирусы человека, животных, членистоногих и растений.

Электронограммы вирионов реовирусов типа I (а) и вирионов орбивирусов — вирус Кемерово (б): хорошо различимы капсомеры (указаны стрелками) внешнего капсидного слоя вирионов реовирусов, капсомеры внешнего капсидного слоя вирионов орбивирусов имеют менее четкие очертания; х 300 000.
Семейство Р. включает 5 родов — Reovirus, Orbivirus, Rotavirus, Phytoreovirus и Fijivirus. Вирионы вирусов, входящих в семейство Р., обладают икосаэдральной структурой, диаметр их колеблется в пределах 60—80 нм, они не имеют липопротеидной оболочки, но содержат двухслойную белковую оболочку, выраженность и строение слоев к-рой варьирует у различных представителей семейства. У вирионов представителей родов Reovirus и Rotavirus в каждом из слоев оболочки выявляются структурные единицы — капсомеры (рис., а), что позволяет относить вирусы этих родов к двухоболочечным (или двухкапсидным). У вирионов представителей родов Orbivirus и Phytoreovirus капсомерная структура внешнего слоя оболочки выражена нечетко (рис., б), поэтому они описываются как однокапсидные вирусы. Структура капсидного слоя вирионов представителей рода Fijivirus изучена недостаточно. Вирионы Р. содержат фрагментированную (10—12 фрагментов) двунитчатую РНК, составляющую 14—22% массы вириона, 6—10 полипептидов, включая транскриптазу и другие ферменты.
В 1978 г. Международный комитет по таксономии вирусов выделил в составе семейства Р. три новых рода — Rotavirus, Phytoreovirus и Fijivirus. К роду Rotavirus (см. Ротавирусы) были отнесены интенсивно изучавшиеся в 1974—1978 гг. возбудители острых гастроэнтеритов человека (см. Ротавирусный гастроэнтерит) и многих видов животных — крупного рогатого скота, белых мышей, морских свинок, овец, коз, свиней, обезьян, лошадей, антилоп, бизонов, оленей, кроликов и собак. Частицы ротавирусов имеют двухкапсидную структуру, отличающуюся от структуры представителей рода Reovirus более четкой выраженностью наружного капсидного слоя, и 11-фрагментную РНК. Роды Phytoreovirus (греч. phyton растение) и Fijivirus (по названию островов Фиджи, где был обнаружен ставший прототипом рода вирус, вызывающий болезнь сахарного тростника) объединяют вирусы, патогенные для растений.
Собственно Р.— представители рода Reovirus — составляют небольшую группу вирусов. Первый представитель рода был выделен и описан Рамос-Альваресом (М. Ramos-Alvarzs) и Сейбином в 1954 г. как один из типов энтеровирусов человека — ECHO-10, однако по значительно большим размерам и ряду других физ.-хим. и биол. свойств отнесен в дальнейшем к самостоятельной группе Р. Многочисленные штаммы представителей рода Reovi-rus, выделенные от людей, обезьян и многих видов низших млекопитающих, оказались принадлежащими к 3 антигенно различным типам — 1, 2 и 3. Штаммы одного типа, выделенные от человека и животных, не отличаются друг от друга. Основные свойства вирионов представителей рода Reovirus соответствуют свойствам, характеризующим семейство Р. Вирионы рода имеют сферическую форму, обладают двумя капсидами; размер вирионов составляет 75— 80 нм. Двунитчатая РНК вирусов рода Reovirus состоит из 10 фрагментов, ее общая мол. масса составляет 14—15 X 106, в составе массы вириона она занимает 14% . Вирион включает 9 полипептидов с мол. массой 34—155 X 103, в состав к-рых входят транскриптаза и нуклеотидфосфогидролаза. Вирусы устойчивы к действию эфира и других растворителей жиров, относительно устойчивы к нагреванию, действию ультрафиолетового света и обычных дезинфицирующих средств, сохраняют активность в широком интервале pH, в т. ч. и при pH 3,0. Подобно энтеровирусам, они приобретают повышенную устойчивость к нагреванию в 1 М р-ре хлорида магния (инфекционный титр вируса не понижается при t °50° в течение 1 часа).
Размножение вирусов рода Reovirus происходит в цитоплазме восприимчивых клеток, где при реови-русной инфекции образуются цитоплазматические околоядерные включения, в к-рых, по-видимому, и осуществляется формирование вирусных частиц. В лабораторных условиях вирусы хорошо размножаются и вызывают цитопатический эффект в культурах клеток многих видов приматов и домашних животных. Для выделения и исследования вирусов применяют культуры клеток почек обезьян макак-резусов или церкопитеков, а также первичные культуры клеток почек эмбриона человека или перевиваемые линии клеток человеческого амниона, HeLa, BS-C-1; в качестве экспериментальных животных наиболее часто используют белых мышей-сосунков.
Вирусы рода Reovirus обладают способностью агглютинировать эритроциты человека, поэтому реакция подавления гемагглютинации находит широкое применение как для идентификации и антигенной группировки Р., так и для диагностических целей (см. Вирусологические исследования, Серологические исследования) .
Р. рода Reovirus трех типов широко распространены во всех районах земного шара, на что указывает обнаружение антител в сыворотках крови людей и животных при массовых обследованиях. Среди млекопитающих Р. имеют широкий спектр хозяев и инфицируют в естественных условиях мышей, кроликов, кошек, собак, свиней, овец, крупный рогатый скот, обезьян макак-резусов, церкопитеков и шимпанзе, несколько видов американских обезьян и других животных. Так как Р. каждого из типов, выделяемые от человека и животных, неразличимы, до сих пор не решен вопрос, существуют ли варианты Р. животных и Р. человека, а если не существуют, то является ли человек первичным хозяином Р. или они должны рассматриваться как вирусы животных, способные инфицировать человека. Инфицированные животные (как и человек) выделяют вирус с фекалиями, к-рые могут служить источником инфекции человека. Р., обильно и длительно выделяющиеся с фекалиями инфицированных коров, могут попадать в молоко. Подобным же образом экскрементами других видов животных, инфицированных Р., могут загрязняться многие пищевые продукты. Сохранению вируса в инфицированных продуктах может способствовать то обстоятельство, что наибольшее распространение реовирусной инфекции наблюдается в осенние и зимние месяцы.
Роль Р. в этиологии ряда заболеваний, с к-рыми они связываются, остается еще неясной. Часто реовирусная инфекция человека протекает бессимптомно. В то же время различные типы Р. были выделены от больных с лихорадочными заболеваниями, экзантематозами, острыми заболеваниями верхних и нижних дыхательных путей (в т. ч. с явлениями интерстициальной пневмонии), жел.-киш. тракта (включая стеаторею) и ц. н. с. (от легких симптомов до энцефалита). Р. удается выделять от больных из носоглоточного отделяемого или из фекалий. В ряде случаев отмечается длительная экскреция Р. с фекалиями и появление в процессе инфекции специфических антител. Однако частое развитие бессимптомной реовирусной инфекции и широкая вариабельность клин, симптомов, развивающихся на фон-е инфицирования организма Р., могут указывать и на то, что Р. в ряде случаев является не этиологическим, а сопутствующим агентом. Экспериментальное инфицирование Р. животных (белых мышей, собак) вызывает в отдельных случаях интерстициальную пневмонию, стеаторею, желтуху. У обезьян макак, зараженных Р. типов 1 и 2, отмечались явления менингита, а при заражении этих животных Р. типа 3 — симптомы гепатита. В то же время убедительные данные о роли Р. в патологии человека отсутствуют.
Библиография: Лабораторная диагностика вирусных и риккетсиозных заболеваний, под ред. Э. Леннета и Н. Шмидт, пер. с англ., с. 285, М., 1974; Феннер Ф. и др. Биология вирусов животных, пер. с англ., т. 1, с. 185, 351, М., 1977; Matthews R. E. F. Classification and nomenclature of viruses, Intervirology, v. 12, p. 130, 1979; Viral and rickettsial infections of man, ed. by F. L. Horsfall a. I. Tamm, p. 569, Philadelphia, 1905.
Читайте также: