
Дифтерийный токсин ингибирует стадию в процессе трансляции
Обновлено: 12.05.2024
Лечение дифтерии. Принципы лечения дифтерии. Профилактика дифтерии. Иммунопрофилактика дифтерии. Дифтерийный анатоксин.
Поскольку патогенез поражений обусловлен действием токсина дифтерии, то основу специфической терапии составляет противодифтерийная лошадиная сыворотка (дифтерийный антитоксин), содержащая не менее 2000 международных антитоксических единиц активности (ME) в 1 мл. Антитоксин вводят внутримышечно или внутривенно в дозах, соответствующих тяжести заболевания (от 20 000 до 100 000 ЕД). Открытие Э. Берингом и Ш. Китазато антитоксических свойств сыворотки иммунных животных явилось одним из важных этапов развития микробиологии, а практическая медицина получила возможность противостоять этой высоколетальной инфекции. Параллельно назначают эффективные антимикробные препараты (аминогликозиды, цефалоспорины), а также проводят симптоматическую терапию. Выписку больных проводят только после двукратного отрицательного результата бактериологического обследования.

Профилактика дифтерии. Иммунопрофилактика дифтерии. Дифтерийный анатоксин
Первоначально развитие заболевания предупреждали введением инактивированного антисывороткой дифтерийного токсина. В настоящее время основу профилактики дифтерии составляют плановая или постэкспозиционная вакцинация.
Для иммунопрофилактики дифтерии применяют дифтерийный анатоксин, разработанный Рамоном.
Дифтерийный анатоксин — токсин, лишенный ядовитых свойств обработкой 0,4% раствором формалина и выдержкой в термостате при температуре 40 °С в течение 30 сут, но сохранивший иммуногенность. Очищенный и концентрированный препарат входит в состав комбинированных вакцин — АКДС, АДС, АДС-М.
• Наличие и содержание AT к дифтерийному токсину определяют в РПГА и РИГА.
• Постинфекциошшй иммунитет нестойкий, поэтому реконвалесценты подлежат вакцинации в общем порядке.
• При выявлении заболевания в детских коллективах контактировавших с заболевшими детьми лиц следует обследовать бактериологически и изолировать от коллектива на 7 сут.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Партнер номинации — компания Cytiva.
Генеральный партнер конкурса — международная инновационная биотехнологическая компания BIOCAD.
Молекулярный аппарат трансляции и его консервативность
Трансляционный цикл на множестве этапов может быть нарушен или замедлен многочисленными и разнородными по химическому строению ингибиторами. Ключевые компоненты трансляционного аппарата клетки — это рибосомы и факторы трансляции (белки, помогающие рибосоме реализовывать разные этапы трансляционного цикла), а также большое количество вспомогательных белков, необходимых для обеспечения синтеза нужными материалами: например, аминоацил-тРНК-синтетазы (АРСазы) — ферменты, соединяющие тРНК с соответствующей ей аминокислотой; или регуляторы биосинтеза белка — компоненты сигнальных путей, связанных с трансляцией. При подборе ингибиторов нельзя не учитывать, что многие из этих молекул есть у всех живых организмов, и некоторые из них довольно консервативны. А значит, одни и те же вещества могут действовать как на про-, так и на эукариот, что не всегда приемлемо.
Каждый живой организм на Земле может быть отнесен к одному из трех доменов жизни: это максимально удаленные друг от друга в эволюционном смысле группы — эукариоты (здесь ищите самые известные царства: животных, растений и грибов), бактерии и археи [2]. Эта удаленность означает, что они давно разошлись (то есть их последний общий предок жил давно [3]), и у представителей групп накопилось множество отличий. Но есть и признаки, по которым представители различных доменов похожи друг на друга. Речь идет об основополагающих признаках: например, способности удваивать ДНК, синтезировать на ее основе РНК и в конце концов получать закодированный в ней белок. Трансляция относится к самым консервативным биологическим процессам.
При ближайшем рассмотрении оказывается, что устройство белоксинтезирующего аппарата довольно схоже у представителей разных доменов. К примеру, основные факторы инициации и элонгации у бактерий имеют гомологов у архей, а рибосомы архей по многим аспектам строения очень напоминают рибосомы эукариот (что даже позволило создать из рибосом дрожжей гибридные рибосомы, в которых одна из субъединица очень напоминала рибосомную субъединицу архей [4]). В пределах этих трех доменов есть отличия, но мы видим, что трансляционная машинерия в целом очень консервативна, и таких отличий не так много. Это является несомненным свидетельством общности (монофилетичности) происхождения жизни.
Ингибиторы трансляции как антибиотики
Чаще всего исследователям приходится подбирать ингибиторы биосинтеза белка у бактерий и заботиться о том, чтобы они не действовали на эукариот, так как это позволяет применять такое вещество в качестве антибиотика. Больше половины используемых сейчас антибиотиков подавляет именно трансляцию, действуя на рибосому , [5]. И здесь на первый план зачастую выходят те тонкие различия, которые все-таки можно отыскать в строении аппарата трансляции у представителей разных доменов.
Трансляционный цикл
Схема может показаться сложной, но в действительности это не так. Попробуйте выхватить взглядом желтую 40S-субъединицу (в свободном от других компонентов трансляции виде ее можно найти рядом со стадией i.1) и пройтись по циклу, как в настольной игре, посмотрев на все, что успевает с ней за это время произойти. Особенно любознательным предлагаем покликать по элементам схемы и прочитать о каждом из этапов дополнительно.
Этапы трансляции
Трансляционный цикл принято подразделять на несколько этапов:
Отдельно происходит активация аминокислот с помощью аминоацил-тРНК-синтетаз (см. e.1 на рисунке 1).
Некоторые трансляционные факторы известны уже очень давно, для них понятна функция, известны ортологи у архей и бактерий (подробнее про гомологичные взаимоотношения генов можно прочитать здесь [7]) и выяснено, являются ли они строго необходимыми для синтеза белка — ведь некоторые вирусные мРНК, например, могут обходиться без целого ряда факторов инициации трансляции [8], [9]. Роль других участников была открыта относительно недавно. Примером таких белков служат факторы DENR и MCTS1 (см. область рециклинга (r) на рисунке 1), изучением которых, кстати, занимается наша лаборатория.
На рисунке 1 (см. s.1, s.2) также изображена часть путей внутриклеточного сигналинга, которые непосредственно регулируют трансляцию. Кроме того, существует еще и сложнейший процесс сборки рибосомы из рибосомной РНК и белков, который протекает по большей части в ядре. Его также можно подавить низкомолекулярными веществами, но о них известно гораздо меньше, а их специфичность часто оставляет желать лучшего.
Типы ингибиторов
С ингибиторами трансляции удобно знакомиться, если классифицировать их согласно мишеням, на которые они воздействуют.
Ингибиторы эукариотической рибосомы
Многие из ингибиторов действуют на рибосомы. При этом они могут быть универсальными (подавлять трансляцию у всех доменов живого), а могут — специфичными (например, к рибосоме эукариот или даже каких-то узких таксономических групп). Специфичность действия, как правило, определяется тонкими различиями в строении места связывания: структурные исследования показывают, что часто достаточно замены одного нуклеотида в рРНК или разницы в единственной аминокислотной позиции рибосомного белка, чтобы конфигурация участка не позволяла ингибитору связаться. Подавляющее большинство ингибиторов рибосомы действует на стадии элонгации, однако существуют и другие типы, перечисленные в таблице 1 (заметьте, что пункты могут перекрываться).
Если классифицировать по сайтам связывания, то выделяют: ингибиторы пептидилтрансферазного центра; ингибиторы транслокации, узнающие E-сайт; ингибиторы, связывающиеся в пептидном тоннеле; ингибиторы, взаимодействующие с декодирующим центром; ингибитор, связывающийся с ГТФаза-активирующим центром (пока достоверно описан только один такой пример).
Пока не так много веществ этого типа допущены до практического применения (самое известное из них — гомохаррингтонин (рис. 2)), но их число постепенно растет. Клинические и доклинические испытания проходили, например, такие ингибиторы пептидилтрансферазного центра, как ликорин, нарциклазин и бруцеантин, а ингибитор транслокации эметин используется как антигельминтное и противомалярийное средство.
Интересные случаи применения
Гомохаррингтонин. Рибосом-направленные ингибиторы элонгации
Гомохаррингтонин — ингибитор пептидилтрансферазного центра в виде полусинтетического аналога омацетаксина мепесукцината (продаваемого под торговой маркой Synribo; рис. 2) активно применяется для лечения хронического миелоидного лейкоза, а также проходит испытания в терапии ряда других онкологических заболеваний. Кроме того, его широко используют в научной работе: его способность прекращать трансляцию новоинициировавших рибосом (при этом не трогая рибосомы, которые уже успели синтезировать несколько пептидных связей) применяется для картирования старт-кодонов в мРНК.
Аталурен и гентамицин. Вещества, вызывающие ошибки декодирования
Рисунок 4. Гентамицин. Пока что препарат прошел клинические испытания только как антибиотик, однако у его производных есть потенциал и в терапии генетических заболеваний. Препарат, продаваемый в аптеках, представляет собой смесь более четырех похожих веществ. Его стоимость куда ниже, чем у аталурена: купить тюбик мази для наружного применения можно за 40 рублей. Однако в нынешнем виде гентамицин нельзя принимать в дозах, достаточных для устранения эффектов нонсенс-мутаций: из-за воздействия на рибосомы митохондрий он имеет побочные эффекты в виде нефро- и ототоксичности.
Чтобы понять, как именно ингибиторы могут подавлять активность рибосомы, полезно вспомнить ее общее строение. Это удобно сделать с помощью интерактивного аплета (см. рис. 5).
Рисунок 5. Пристальный взгляд на эукариотическую рибосому. В верхней кнопочной панели можно изучить структурные элементы рибосомы. Для того чтобы узнать о структурном элементе или сайте связывания, наведите курсор на кнопку. В нижней панели можно найти распространенные ингибиторы, которые действуют на разные части эукариотической рибосомы, визуализированные в предыдущем разделе. В виде на лиганд взаимодействующие основания выделены оранжевым. Встроенный в статью этот апплет очень маленький, поэтому рекомендуем открыть полноэкранную версию.
- Чтобы увидеть лиганд-белковые взаимодействия, можно кликнуть на лиганд в режиме его просмотра.
- Из-за большого размера структур и их сложности для просмотра рекомендуется пользоваться компьютером со стабильным подключением.
- При переходе между видом рибосомы и видом на лиганд структура перезагружается.
- К сожалению, выделение структурных элементов из прошлого раздела в структурах с ингибиторами не поддерживается.
- В будущем рисунок будет совершенствоваться, при клике на хищную рибосому читатель может найти более совершенную версию, которая периодически обновляется.
Для показа структурных элементов использована структура дрожжевой рибосомы, полученная методом криоэлектронной микроскопии группой Р. Бекманна (6SNT). Для визуализации используется веб-плагин MolStar.
Ингибиторы трансляционных факторов
Трансляционные факторы — белки, помогающие рибосоме реализовывать разные этапы трансляционного цикла. Чаще всего эти факторы жизненно необходимы для нормального синтеза белка в клетках или митохондриях/пластидах, которые, как правило, тоже имеют свой трансляционный аппарат.
Эта группа веществ взаимодействует с факторами в растворе, блокируя их активность или препятствуя присоединению к рибосоме (во втором случае они могут контактировать не только с фактором, но и с рибосомой, что приводит к неоднозначности в классификации: иногда их относят к ингибиторам рибосомы).
Ингибиторы факторов инициации трансляции
Инициация трансляции в эукариотических клетках устроена сложнее, чем у бактерий: факторов инициации здесь больше, и многие из них являются эукариот-специфичными. Поэтому и набор мишеней для ингибиторов довольно разнообразен. Со всем многообразием факторов инициации и их набором у трех доменов живого удобно ознакомится в таблице 2.
* — IF3 бактерий не гомологичен a/eIF1, но очень похож на него по пространственной структуре.
** — Каталитические γ- и ε-субъединицы у архей отсутствуют, поэтому вопрос о наличии целостного aIF2B и даже об участии остальных субъединиц в трансляции неясен: на этот счет в научной литературе до сих пор ведутся споры.
Ингибиторы факторов элонгации
На данный момент известно большое количество ингибиторов фактора eEF2 (транслоказы), однако некоторые вещества действуют и на eEF1A, доставляющий аминоацил-тРНК. Бóльшая часть и тех, и других препятствует диссоциации факторов с рибосомы после гидролиза ГТФ, что блокирует цикл элонгации. Некоторые их этих ингибиторов (сордарин и его производные) обладают специфичностью к eEF2 грибов, но не связываются с фактором из человеческих клеток, что позволяет использовать их в качестве противогрибковых средств. Среди известных веществ из этой категории нельзя не упомянуть плитидепсин (аплидин), который, согласно работе, недавно опубликованной в журнале Science, показывает хорошие результаты в борьбе с вирусом SARS-CoV-2 [15].
Ингибиторы аминоацил-тРНК-синтетаз
Помимо рибосом и фактор-направленных ингибиторов, специфичное влияние на биосинтез белка могут оказывать вещества, блокирующие вспомогательные компоненты трансляционной машинерии — в первую очередь ингибиторы аминоацил-тРНК-синтетаз (АРСаз). Фундаментальной основой трансляции является генетический код (соответствие между триплетами нуклеотидов и аминокислотами). Осуществляют это соответствие не столько рибосома (хотя в ее функции, разумеется, входит контроль правильного декодирования триплетов — то есть соответствия тРНК кодону в мРНК), сколько АРСазы, навешивающие аминокислотный остаток на нужную тРНК. Из примерно 20 аминоацил-тРНК-синтетаз эукариот (точное их число может варьировать от вида к виду) специфичные ингибиторы известны как минимум для семи.
При ингибировании АРСаз в клетке уменьшается концентрация Аа-тРНК, и темпы трансляции снижаются. От этого сильнее всего страдают клетки с высокой интенсивностью белкового синтеза — к примеру, иммунные и раковые. Поэтому ингибиторы АРСаз имеют важное медицинское значение: они обладают иммуносупрессорным эффектом и применяются при лечении опухолей. Их также используют в качестве фунгицидов и антималярийных препаратов. Большинство ингибиторов этого типа высокоспецифичны по отношению к конкретным АРСазам, однако вещество пурпуромицин препятствует присоединению аминокислотного остатка к любой тРНК. На рисунке 1 ингибиторы АРСаз обозначены e.1.
Ингибиторы сигнальных путей, связанных с регуляцией трансляции
Существует несколько сигнальных путей, которые тонко регулируют трансляцию на разных уровнях. Эти пути образуют сложную сеть и на схеме отображены лишь частично. Ключевые регуляторные каскады, контролирующие эффективность трансляции в клетке, — это пути PI3K/AKT/mTOR и MAPK/ERK/Mnk, а также набор киназ α-субъединицы фактора eIF2. Ингибиторы mTOR представляют особую ценность из-за выраженных комплексных эффектов на организм, у них множество применений в медицине: к примеру, они используются при лечении рака или как иммуносупрессоры при трансплантациях органов. Самый известный пример лекарств последнего типа — это рапамицин (сиролимус), который ингибирует один из двух комплексов киназы mTOR (mTORC1). С этим лекарством, а также с другими ингибиторами mTOR связаны и надежды на замедление темпов старения: показано, что некоторые из этих веществ ощутимо продлевают период здоровой жизни у мышей [16]. На рисунке 1: s.1–s.9.
Немного о научной группе
Наша лаборатория расположена в отделе взаимодействия вирусов с клеткой НИИ физико-химической биологии имени А.Н. Белозерского МГУ. Основные направления исследований — изучение механизмов биосинтеза белка, молекулярная вирусология и анализ возрастных изменений в живых организмах на молекулярном и клеточном уровне. В отделе работают несколько научных сотрудников, аспирантов и множество студентов, руководитель отдела — к.б.н. Сергей Дмитриев. В исследованиях мы применяем широкий арсенал методов молекулярной биологии, генной инженерии, биохимии, системной биологии и биоинформатики. Со списком публикаций отдела можно ознакомиться на сайте МГУ.
Рисунок 6. Структуры из нашей лаборатории. А пока можете посмотреть на структуры белков, которыми занимается лаборатория и которые были получены нашими партнерами с помощью рентгеноструктурного анализа и криоэлектронной микроскопии. Также вы можете открыть полноэкранную версию апплета.
Токсинообразование дифтерийной палочкой. Токсины дифтерии. Дифтерийный токсин. Способность к токсинообразованию дифтерийной палочки. Активность токсина дифтерии.
Corynebacterium diphtheriae продуцирует мощный экзотоксин — основной фактор патогенности. Нетоксигенные штаммы дифтерии не вызывают развитие заболевания. Б чистом виде токсин впервые получили ; Э. Ру и А. Иерсен (1888), что явилось решающим моментом для установления этиологической роли микроорганизма.
Токсин дифтерии проявляет все свойства экзотоксина (термолабильный, высокотоксичный, иммуногенный белок, нейтрализуемый антитоксической сывороткой). Нативный токсин — полипептид с Мг около 72 000; его образуют фрагменты А (проявляет ферментативную активность) и В (взаимодействует с клеточными рецепторами, облегчая проникновение фрагмента А). Клетки всех чувствительных организмов способны рецептировать В-фрагмент и поглощать молекулу посредством эндоцитоза. В кислой среде эндосом (фаголизосом) дисульфидные связи, объединяющие оба компонента, разрушаются, фрагмент В взаимодействует с мембраной эндосомы, облегчая проникновение фрагмента А в цитоплазму. Последний устойчив к денатурации и длительно сохраняется в цитозоле.
Механизм цитотоксического действия токсина дифтерии связан с модификацией белков через АТФ-рибозилирование. Подобным свойством обладают многие токсины, но лишь дифтерийный токсин и токсин A Pseudomonas aeruginosa имеют специфичную мишень — фактор элонгации 2 — трансферазу, ответственную за наращивание (элонгацию) полипептидной цепи на рибосоме.

Токсины дифтерии. Дифтерийный токсин
Дифтерийный токсин катализирует перенос АТФ-рибозы от цитоплазматического никотин-амиддинуклеотида (НАД) к фактору элонгации 2, приводя к АТФ-рибозилированию гистидиновых остатков в молекуле фактора с необратимым блокированием элонгации полипептидной цепи (то есть любого белкового синтеза). Немодифицированный фактор элонгации 2 образует комплекс с ГТФ и тРНК, связывающийся с мРНК в эукариотических клетках, после чего возможно встраивание аминокислот в синтезируемую белковую молекулу.
Токсин дифтерии ингибирует белковый синтез, в том числе и в миокарде, приводя к структурным и функциональным нарушениям, способным вызвать смерть больного. Результат действия токсина дифтерии на нервную ткань — демиелинизация нервных волокон, часто приводящая к параличам и парезам.

Способность к токсинообразованию дифтерийной палочки
Способность к токсинообразованию проявляют лишь лизогенные штаммы Corynebacterium diphtheriae, инфицированные бактериофагом (р-фаг), несущим ген fox, кодирующий структуру токсина дифтерии. Образование последнего наиболее выражено при вступлении бактериальной популяции в стадию отмирания. Переход умеренного фага в литическую форму мало влияет на синтез токсина.
Активность токсина дифтерии
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Один из путей выяснения тонких молекулярных механизмов синтеза нуклеиновых кислот и белков в клетках – использование таких лекарственных препаратов, которые могли бы избирательно тормозить эти процессы у бактерий, не влияя на клетки организма человека. Некоторые препараты, действительно, оказывают такое избирательное действие, взаимодействуя с белками рибосом прокариот и выключая бактериальный синтез белка. Однако многие из них являются токсичными и для человека. В настоящее время в медицинской практике применяются многие антибиотики, часть из которых будет рассмотрена с целью выяснения молекулярного механизма их действия на ключевые химические реакции синтеза белка и нуклеиновых кислот.
Один из мощных ингибиторов белкового синтеза – пуромицин. Он представляет собой аналог концевого участка аминоацил-тРНК адениловой кислоты и поэтому легко взаимодействует с А-центром пептидил-тРНК с образованием пептидил-пуромицина :
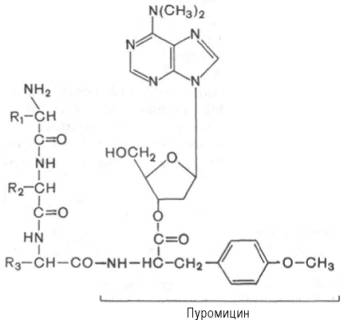
Пептидил-пуромицин не несет на себе триплета антикодона и поэтому тормозит элонгацию пептидной цепи, вызывая обрыв реакции, т.е. преждевременную терминацию синтеза белка. При помощи пуромицина было доказано, например, что гормональный эффект в ряде случаев зависит от синтеза белка de novo. Укажем также, что пуромицин оказывает тормозящее действие на синтез белка как у прокариот, так и у эукариот.
Белковый синтез тормозится актиномицином D, обладающим противоопухолевым эффектом, однако вследствие высокой токсичности препарат применяется редко. Он тормозит синтез всех типов клеточной РНК, особенно мРНК. Данное свойство объясняется тормозящим влиянием актиномицина D на ДНК-зависимую РНК-полимеразу, поскольку он связывается с остатками дезоксигуанозина цепи ДНК, выключая матричную функцию последней; это дает основание считать, что актиномицин D ин-гибирует транскрипцию ДНК.
Другим антибиотиком, также тормозящим синтез клеточной РНК, является используемый при лечении туберкулеза рифамицин. Этот препарат тормозит ДНК-зависимую РНК-полимеразу, связываясь с ферментом. Наиболее чувствительной к нему оказалась бактериальная РНК-полиме-раза. На организм животных этот антибиотик оказывает незначительное влияние. По механизму действия он резко отличается от актиномицина D. Следует указать, кроме того, на недавно открытое противовирусное действие рифамицина; в частности, он успешно используется при лечении трахомы, которая вызывается ДНК-содержащим вирусом. Это дает основание предположить, что данный антибиотик найдет применение в клинической онкологии при лечении опухолей, вызываемых вирусами.
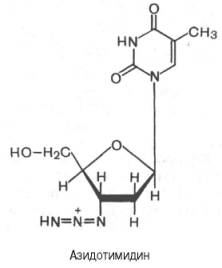
Одним из мощных ингибиторов синтеза вирусной РНК оказался азидотимидин (3'-азидо-2',3'-дидезокситимидин), синтезированный еще в 1964 г. в надежде на его противоопухолевый эффект. Было показано, что вирус иммунодефицита человека (ВИЧ) содержит РНК-й геном, в составе которого имеются как стандартные гены ретровирусов, так и необычные небольшие гены со множеством функций. Последние, в частности, подвержены мутациям с высокой скоростью вследствие низкой точности репликации, вызванной свойствами обратной транскриптазы. Эта вирусная обратная транскриптаза иммунодефицита человека оказалась наделенной значительно большим сродством к азидотимидину, чем к природному дезокситимидинтрифосфату (dTТФ). Азидотимидин конкурентно тормозит связывание dTТФ, вызывая тем самым терминацию (окончание) синтеза вирусной РНК.
Выяснены некоторые детали механизма действия ряда других антибиотиков, используемых при лечении тифозных инфекций. Так, хлорам-феникол оказывает ингибирующее влияние на пептидилтрансферазную реакцию (на стадии элонгации) синтеза белка в 70S рибосоме бактерий; на этот процесс в 80S рибосоме он не действует. Тормозит синтез белка в 80S рибосоме (без поражения процесса в 70S рибосоме) циклогексимид – специфический ингибитор транслоказы.
Весьма интересен молекулярный механизм действия дифтерийного токсина. Он оказался наделенным способностью катализировать реакцию АДФ-рибозилирования фактора элонгации эукариот (eEF-2), выключая тем самым его из участия в синтезе белка. Резистентность многих животных к дифтерийному токсину, вероятнее всего, обусловлена трудностью или полным отсутствием проникновения (транспорта) токсина через мембрану клеток.
Противотуберкулезные и антибактериальные антибиотики, в частности стрептомицин и неомицин, действуют на белоксинтезирующий аппарат чувствительных к ним штаммов бактерий. Было высказано предположение, что эти антибиотики обусловливают ошибки в трансляции мРНК, приводящие к нарушению соответствия между кодонами и включаемыми аминокислотами: например, кодон УУУ вместо фенилаланина начинает кодировать лейцин, в результате чего образуется аномальный белок, что приводит к гибели бактерий.
Широко применяемые в клинике тетрациклины также оказались ингибиторами синтеза белка в 70S рибосоме (меньше тормозится синтез в 80S рибосоме). Они легко проникают через клеточную мембрану. Считают, что тетрациклины тормозят связывание аминоацил-тРНК с аминоацильным центром в 50S рибосоме. Возможно, что тетрациклины химически связываются с этим центром, выключая тем самым одну из ведущих стадий процесса трансляции.
Пенициллины не являются истинными ингибиторами синтеза белка, однако их антибактериальный эффект связан с торможением синтеза гексапептидов, входящих в состав клеточной стенки. Механизм их синтеза отличается от рибосомного механизма синтеза белка. Эритромицин и олеандомицин тормозят активность транслоказы в процессе трансляции, подобно циклогексимиду, исключительно в 80S рибосомах, т.е. тормозят синтез белка в клетках животных.
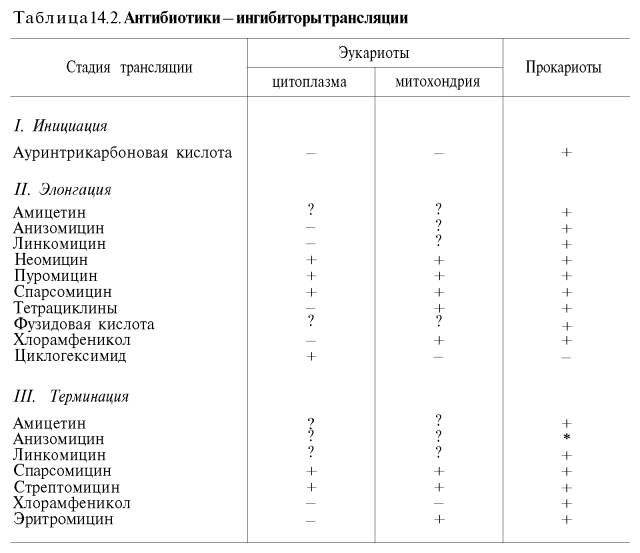
Полученные к настоящему времени данные о механизме действия антибиотиков на синтез белка с учетом стадии и топографии процесса трансляции суммированы в табл. 14.2 (по Харперу с небольшими изменениями).
Следует еще раз подчеркнуть, что нарушение или выпадение любого звена, участвующего в синтезе белка, почти всегда приводит к развитию патологии, причем клинические проявления болезни будут определяться природой и функцией белка, синтез которого оказывается нарушенным (структурный или функциональный белок). Иногда синтезируются так называемые аномальные белки как результат действия мутагенных факторов и соответственно изменения генетического кода (например, гемоглобин при серповидно-клеточной анемии). Последствия этих нарушений могут выражаться в развитии самых разнообразных синдромов или заканчиваться летально.
Следует отметить, однако, что организм располагает мощными механизмами защиты. Подобные изменения генетического аппарата быстро распознаются специфическими ферментами – рестриктазами, измененные последовательности вырезаются и вновь замещаются соответствующими нуклеотидами при участии полимераз и лигаз.

Дифтерия - это острое инфекционное заболевание, опасное для жизни. Наиболее восприимчивы к нему дети в возрасте 3–7 лет, в последние годы увеличилась заболеваемость подростков и взрослых.
Дифтерия – это острое инфекционное заболевание, опасное для жизни. Оно протекает в виде острого воспаления верхних дыхательных путей, преимущественно глотки (примерно 90% случаев), носа, кожи в местах ее повреждения, глаз или половых органов.
Основную угрозу, однако, представляет не воспаление, а отравление токсином, который вырабатывает бактерия – возбудитель заболевания, при этом преимущественно поражаются сердечно-сосудистая и нервная системы.
Возбудитель дифтерии и пути заражения
Возбудителем дифтерии являются Corynebacterium diphteriae – грамположительные бактерии в виде палочек с характерными колбовидными утолщениями на концах, которые в мазках располагаются попарно, под углом в виде римской цифры V по отношению друг к другу. Дифтерийные палочки в процессе жизнедеятельности выделяют дифтерийный токсин, фермент нейраминидазу и другие биохимически активные соединения.
Синтез дифтерийного токсина микробными клетками контролируется специальным геном tox. Бактерии могут терять его в процессе жизнедеятельности, утрачивая вместе с ним и свою способность вырабатывать токсин (токсигенность). И, наоборот, изначально нетоксигенные штаммы могут приобретать патогенные свойства, к счастью, это случается крайне редко.
Заболевание передается воздушно-капельным путем от больных дифтерией или от здоровых носителей инфекции, значительно реже – через предметы обихода.
Группа риска
Наиболее восприимчивы к заражению дифтерией дети в возрасте 3–7 лет, но в последние годы увеличилась заболеваемость подростков и взрослых.
Источником инфекции являются больные люди или здоровые носители токсигенных бактерий. Наиболее заразны страдающие дифтерией зева, носа и гортани, так как они активно выделяют возбудителей заболевания с выдыхаемым воздухом. Больные дифтерией глаз, кожи могут распространять инфекцию контактным путем (руки, предметы быта). Здоровые носители бактерий гораздо менее заразны, но отсутствие у них каких-либо внешних признаков их состояния не позволяет контролировать распространение ими инфекции, ведь выявить их можно только случайно в ходе массовых диспансерных обследований. В результате большинство случаев заражения дифтерией обусловлено контактом со здоровыми носителями дифтерийной палочки.
Инкубационный период (время от момента заражения до появления первых признаков заболевания) составляет 2–10 дней.
Дифтерийный токсин
Токсин, вырабатываемый дифтерийной палочкой, состоит из нескольких компонентов. Один из них – фермент гиалуронидаза разрушает гиалуроновую кислоту капилляров и увеличивает их проницаемость, что приводит к выходу из сосудов и пропитыванию окружающих тканей плазмой крови с отложением белка фибриногена. Второй компонент – некротоксин разрушает клетки эпителия с выделением из них фермента тромбокиназы. Тромбокиназа способствует превращению фибриногена в фибрин и образованию на поверхности тканей фибринной пленки. При действии дифтерийного токсина на небные миндалины, которые покрыты несколькими слоями эпителиальных клеток, формируется фибиринная пленка, проникающая вглубь эпителия миндалин и плотно спаянная с ним.
Третий (основной) компонент – собственно токсин способен блокировать процессы клеточного дыхания и синтеза белковых молекул. Наиболее чувствительными к его действию являются капилляры, клетки миокарда и нервные клетки. В результате развивается дистрофия миокарда и инфекционно-токсический миокардит, повреждение капилляров приводит к инфекционно-токсическому шоку, повреждение шванновских клеток (вспомогательные клетки нервной ткани) приводит к демиелинизации нервных волокон (разрушение электроизолирующего слоя миелина с нарушением проведения нервных импульсов по нервным волокнам). Кроме того, дифтерийный токсин вызывает общую интоксикацию организма.
Симптомы и течение
Дифтерия зева обычно начинается с небольшого повышения температуры, незначительной болезненности при глотании, покраснения и отечности миндалин, формированию на них специфического пленчатого налета, увеличения передних верхних шейных лимфатических узлов. Цвет пленок – обычно белый в первые 2–3 дня заболевания, но потом приобретает серый или желтовато-серый цвет. Примерно через неделю заболевание, или заканчивается выздоровлением (легкая форма, как правило, у привитых от дифтерии), или переходит в более тяжелую токсическую форму, обусловленную системным действием дифтерийного токсина.
Токсическая форма дифтерии всегда протекает очень тяжело. Она характеризуется очень высокой температурой тела (39,5-41,0°С), сильными головными болями, сонливостью, апатией. Кожа становится бледной, во рту отмечается сухость, у детей возможна многократная рвота и боль в животе. Отек миндалин становится резко выраженным, может приводить к полному закрытию входа в глотку, распространяется на мягкое и твердое небо, часто также на носоглотку, дыхание затрудняется, голос часто становится гнусавым. Налет распространяется на все ткани ротоглотки. Классическим признаком токсической формы дифтерии зева является отек подкожной клетчатки в области шеи, а иногда и грудной клетки, в результате чего кожа приобретает студнеобразную консистенцию. Передние верхние шейные лимфатические узлы значительно увеличены и болезненны.
Дифтерия носа протекает на фоне нормальной или слегка повышенной температуры тела, интоксикация отсутствует. Из носовых ходов видно серозно-гнойное или кровянисто-гнойное отделяемое. На крыльях носа, щеках, лбу и подбородке появляются участки мокнутия, а затем сухие корочки. Внутри носа видны пленчатые налеты. Патологический процесс может также поражать придаточные пазухи носа. При токсической форме наблюдается отек подкожной клетчатки щек и шеи.
Дифтерия глаза протекает как банальный конъюнктивит и характеризуется умеренной гиперемией и отеком конъюнктивы века, небольшим количеством серозно-гнойного отделяемого из конъюнктивального мешка (катаральная форма). Пленчатая форма проявляется выраженным отеком век, наличием на их конъюнктиве трудно снимаемых пленок серовато-белого цвета. Токсическая форма также сопровождается отеком клетчатки вокруг глазницы.
Дифтерия кожи приводитк длительному не заживлению любых повреждений кожи, гиперемии, на коже присутствует грязно-серый налет, отмечается плотная инфильтрация окружающей кожи.
Диагностика
Диагностику дифтерии проводят на основании данных осмотра больного и результатов анализов. При осмотре за диагноз дифтерия говорят следующие признаки: наличие характерных пленок, а также затруднение дыхания и свистящий шум на вдохе, не характерные для ангины, лающий кашель. Диагноз дифтерии по характерным клиническим признакам при заболевании, протекающем в легкой форме, поставить сложнее.
Общий анализ крови – признаки острого воспалительного процесса.
Исследования мазка под микроскопом (бактериоскопия) – выявление имеющих характерный вид бактерий Corynebacterium diphteriae.
Бактериологическое исследование – посев биологического материала на специальную питательную среду и культивирование колоний микроорганизмов.
Определение уровня (титра) антитоксических антител (высокий титр – 0,05 МЕ/мл и выше позволяет исключить дифтерию).
Серологическое исследование – определение специфических антител в сыворотке крови с помощью методов РПГА, ИФА и др.
Дифтерию зева нужно дифференцировать от острого тонзиллита (фолликулярная и лакунарная формы), ангины Симановского-Венсана (грибковое поражение), сифилитической ангины, ложнопленочной ангины при инфекционном мононуклеозе, паратонзиллярным абсцессом, эпидемическим паротитом, лейкозом. У детей необходимо отвести диагноз ложного крупа.
Лечение
Всех заболевших дифтерией, вне зависимости от тяжести состояния, необходимо госпитализировать в инфекционную больницу.
Лечение состоит в следующем:
Диета – витаминизированная, калорийная, прошедшая тщательную кулинарную обработку еда.
Этиотропная терапия (то есть направленная на устранение причины заболевания) – введение противодифтерийной сыворотки (ПДС), доза и количество раз введения зависят от тяжести и формы заболевания. При легкой форме ПДС вводится однократно внутримышечно в дозе 20–40 тыс. МЕ, при среднетяжелой форме – 50–80 тыс. МЕ однократно или, при необходимости, повторно в той же дозе через 24 часа. При лечении тяжелой формы заболевания суммарная доза увеличивается до 90–120 тыс. МЕ или даже до 150 тыс. МЕ (инфекционно-токсический шок, ДВС-синдром). При этом 2/3 дозы вводится сразу, и в течение первых суток госпитализации должно быть введено 3/4 суммарной дозы.
Антибиотики: при легких формах – эритромицин, рифампицин внутрь, при среднетяжелых и тяжелых формах – инъекционное введение пенициллинов или цефалоспоринов. Продолжительность курса – 10–14 дней. Антибиотики не влияют на дифтерийный токсин, но уменьшают количество бактерий, его вырабатывающих.
Местное лечение – полоскания и орошения дезинфицирующими растворами.
Дезинтоксикационная терапия – глюкозо-солевыми растворами с учетом суточной потребности в жидкости и ее потерь (среднетяжелая и тяжелая форма).
Глюкокортикостероиды – при среднетяжелой и тяжелой формах.
Лечение бактерионосителей проводят антибиотиками: тетрациклины (дети старше 9 лет), эритромицин, цефалоспорины на фоне общеукрепляющей терапии и устранения хронических очагов инфекции.
Осложнения
Среди наиболее серьезных осложненийдифтерии на сердечно-сосудистую систему можно выделить миокардиты, нарушения сердечного ритма.
Неврологические осложнения дифтерии обусловлены поражением различных черепных и периферических нервов и проявляются параличом аккомодации, косоглазием, парезами конечностей, а в более тяжелых случаях параличом дыхательных мышц и мышц диафрагмы.
Вторичными осложнениями дифтерии являются такие тяжелые патологические состояния как острые нарушения мозгового кровообращения (тромбозы, эмболия), метаболическая энцефалопатия, отек головного мозга, токсические поражения почек, дифтерийный гепатит, а также инфекционно-токсический шок и ДВС-синдром (синдром дессиминированного внутрисосудистого свертывания – тяжелое нарушение системы свертывания крови). Токсическая форма дифтерии может приводить к острой почечной, сердечно-сосудистой, дыхательной или полиорганнной недостаточности.
Неспецифическими осложнениями дифтерии являются паратонзиллярный абсцесс, отит, пневмония.
Вакцинация
Вакцинация от дифтерии проводится анатоксином, то есть инактивированным токсином. В ответ на его введение в организме образуются антитела не к Corynebacterium diphteriae, а к дифтерийному токсину.
Согласно российскому прививочному календарю, вакцинация детей в возрасте до года проводится в 3, 4–5 и 6 месяцев. Первая ревакцинация проводится в 18 месяцев, вторая – в 7 лет, третья – в 14. Взрослые должны ревакцинироваться от столбняка и дифтерии каждые 10 лет.
Читайте также: