
Формы вирусных инфекций продуктивная и персистирующая
Обновлено: 05.05.2024
Продуктивная вирусная инфекция с образованием дочерних популяций и характерными клиническими проявлениями возможна лишь при наличии в заражённом организме чувствительных клеток, в которых осуществляется репродуктивный цикл возбудителя. Например, возбудитель полиомиелита может реплицировать только в клетках ЖКТ и ЦНС приматов и человека.
Абортивная инфекция развивается при проникновении возбудителя в нечувствительные клетки (например, при попадании вируса лейкоза коров в организм человека) либо в клетки, не способные обеспечить полный репродуктивный цикл (например, находящиеся в стадии клеточного цикла G0). Способность клеток к поддержанию вирусспецифических репродуктивных процессов также подавляет ИФН, противовирусный эффект которого направлен против самых различных вирусов.
Персистирующая вирусная инфекция возникает при таком взаимодействии между вирусом и заражённой клеткой, когда в последней продолжается выполнение собственных клеточных функций. Если заражённые клетки делятся, образуется инфицированный клон. Таким образом, увеличение числа заражённых клеток способствует увеличению общей популяции возбудителя в организме. Тем не менее персистирующие вирусные инфекции обычно нарушают функции клеток, что в конце концов приводит к клиническим проявлениям. У человека развитие персисти-рующих инфекций в определённой степени зависит от возраста. Например, внутриутробное заражение вирусом коревой краснухи или цитомегаловирусом (ЦМВ) приводит к ограниченному по времени персистированию возбудителя. Появление симптоматики связано с возможностью плода развивать иммунные реакции на инфекционный агент.
Латентная (скрытая) вирусная инфекция. В то время как персистирующие инфекции сопровождаются постоянным высвобождением дочерних вирусных популяций, при латентных поражениях они образуются спорадически. Репродуктивный цикл подобных возбудителей резко замедляется на поздних стадиях и активируется под влиянием различных факторов. Латентные инфекции характерны для большинства герпесвирусов, вызывающих рецидивирующие и обычно не прогрессирующие заболевания.
Дремлющая (криптогенная) вирусная инфекция — форма проявления вирусной инфекции при которой возбудитель в неактивном состоянии находится в отдельных очагах (например, в нервных ганглиях). Клинически инфекция проявляется лишь при резком ослаблении защитных сил организма. Например, вирус герпеса 3 типа, вызывающий при первичном заражении ветряную оспу, пожизненно сохраняется в организме. Рецидив заболевания в форме опоясывающего лишая возможен лишь при нарушениях иммунного статуса (наиболее часто в пожилом возрасте).
Медленные вирусная инфекции характеризуются длительным инкубационным периодом (месяцы и годы), в течение которого возбудитель размножается, вызывая всё более явные повреждения тканей. Первоначально возбудитель размножается в ограниченной группе клеток, но постепенно инфицирует всё большее их число. Заболевания заканчиваются развитием тяжёлых поражений и смертью больного. К медленным вирусным инфекциям относят подострый склерозирующий панэнцефалит, ВИЧ-инфекцию и др.
Вирусные инфекции. Особенности патогенеза вирусных инфекций. Основные этапы патогенеза вирусных инфекций.
В основе патогенеза вирусных инфекций лежит взаимодействие генома вируса с генетическим аппаратом чувствительной клетки. Исключая поражения, вызываемые вирусами, распространяющимися по нервной ткани, патогенез вирусных инфекций сопровождает вирусемия (виремия)— циркуляция возбудителя в крови. В кровоток возбудитель проникает прямым путем или из лимфатической системы. Многие вирусы (например, ВИЧ, вирусы гриппа, кори, герпеса) поражают иммунокомпетентные клетки, что чаще проявляется в нарушении функций и уменьшении числа Т-хелперов, увеличении содержания и активации Т-супрессоров или В-клеток. Некоторые возбудители образуют внутриядерные или цитоплазматические тельца включений (например, тельца Бабеша~Нёгри). имеющие диагностическое значение.
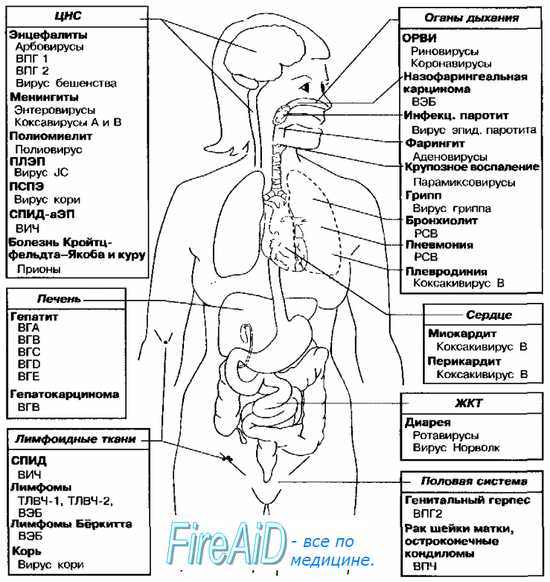
Рис. 5-9. Органы-мишени наиболее распространённых вирусных инфекций человека. ВЭБ — вирус Эпстайна-Барр, ВГА — вирус гепатита А, ВГВ — вирус гепатита В, ВГС — вирус гепатита С, BID — вирус гепатита D, ВГЕ — вирус гепатита Е, ВПЧ — вирус папилломы человека, ВПГ — вирус простого герпеса, ТЛВЧ — Т-пимфотропный вирус человека, ВИЧ — вирус иммунодефицита человека, ПЛЭП — прогрессирующая лейкоэнцефалопатия, РСВ — респираторно-синцитиальный вирус, ПСПЭ — подострый склерозирующий панэнцефалит, ОРВИ — острая респираторная вирусная инфекция, СПИД-аЭП — СПИД-ассоциированная энцефалопатия.
Основные этапы патогенеза вирусных инфекций
Проникновение вируса в организм. Основные входные ворота для возбудителей вирусных инфекции человека — дыхательные пути и ЖКТ, реже — кожные покровы. В некоторых случаях развиваются локальные поражения, но чаще в месте проникновения не возникает каких-либо проявлений или они носят стёртый характер, а возбудитель мигрирует в чувствительные ткани. Распространение возбудителя в организме может носить локальный или системный характер.
Локальные поражения вирусами типичны для возбудителей респираторных и кишечных инфекций, а также для некоторых кожных заболеваний. Продолжительность инкубационного периода большинства подобных инфекций составляет 2-3 сут. Первичную репликацию часто сопровождает вирусемия. Она обычно протекает бессимптомно или по типу продромальных явлений, но может возникать и на фоне выраженной клинической картины, не вызывая развития дополнительной симптоматики. Для подобных заболеваний характерно повторное заражение, так как циркулирующие AT не проявляют протективный эффект, а секреторный иммуноглобулин А (IgA) оказывает лишь кратковременное нейтрализующее действие на слизистой оболочке. Системные поражения. Из места проникновения возбудители попадают в кровоток, вызывая вирусемию, и постепенно фиксируются в чувствительных тканях. Первичное распространение обычно вызывает продромальные явления. Поскольку вирусемия предшествует поражению чувствительных тканей, то продолжительность инкубационного и продромального периодов подобных инфекций могут увеличиваться до 2-3 нед. Вирусемия при системных инфекциях обычно носит двухэтапный характер. Первый этап заканчивается поглощением циркулирующих вирусов клетками ретикулоэндотелиальной системы.
В дальнейшем возможно несколько вариантов:
• полная элиминация возбудителя (абортивная инфекция);
• размножение вирусов в фагоцитах с последующим выходом и развитием выраженной вторичной вирусемии, сопровождающейся появлением характерных клинических признаков заболевания (например, энцефалитов);
• некоторые вирусы (например, вирус гепатита В, пикорна- и тогавирусы) слабо поглощаются фагоцитами и могут циркулировать в крови в свободном состоянии, а возбудители колорадской клещевой лихорадки и лихорадки долины Рифт внедряются в эритроциты.
Основные органы-мишени наиболее распространённых вирусных инфекций представлены на рис. 5-9. Многие из указанных на рисунке возбудителей могут поражать, кроме названных, и другие ткани (так, полиовирусы способны вызывать поражения ЖКТ, а вирус эпидемического паротита обладает тропностью к эпителию извитых канальцев яичек).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
В заражённых вирусом клетках возможны патологические проявления разностороннего характера.
Морфология патологических эффектов. Зараженные вирусом клетки. Основные реакции зараженных вирусом клеток.
Характерные проявления взаимодействий между вирусом и чувствительными клетками-видимые поражения заражённых клеток вплоть до их гибели, а также присутствие возбудителей в исследуемом материале.
Альтерация и воспаление зараженных вирусом тканей. При вирусных инфекциях на первый план выступает картина повреждения клеток и воспалительных изменений тканей, при различных инфекциях их соотношение и выраженность варьируют. В противоположность бактериальным инфекциям (где доминируют полиморфноядерные лейкоциты), при вирусных поражениях среди клеточных элементов воспалительных реакций доминируют мононуклеары (лимфо- и моноциты). На этапах, предшествующих разрушению клеток, можно визуально наблюдать их дегенеративные и некротические изменения.
Тельца включений зараженных вирусом клеток. Микроскопия заражённых клеток часто позволяет выявить тельца включений — характерный, но не абсолютный признак вирусных поражений. Тельца значительно крупнее, чем отдельные вирионы, и часто окрашиваются кислыми красителями (например, эозином).
• При заражении клеток ДНК-содержащими вирусами тельца включений располагаются в ядре; исключение — тельца включений поксвирусов (тельца Гварнери).
• При заражении клеток РНК-содержашими вирусами тельца включений располагаются в цитоплазме (например, тельца Бабеша-Нёгри, выявляемые в цитоплазме клеток головного мозга при бешенстве).
Причины гибели зараженных вирусом клеток. Размножаясь в клетке, вирусы индуцируют синтез вирусспецифических белков, в той или иной степени подавляющих метаболизм клетки. Нарушение синтеза макромолекул вызвано нарушением трансляции клеточной мРНК. Среди РНК-геномных вирусов наиболее быстрое и глубокое подавление макромолекулярных синтезов в клетке вызывают пикорнавирусы, среди ДНК-геномных — покс- и герпесвирусы. Действие указанных вирусов реализуется на ранних этапах (до появления морфологических признаков цитопатического эффекта). Ингибирование синтеза РНК и ДНК обычно вторично по отношению к воздействию на белки, контролирующие экспрессию генов и пролиферацию клетки. Значительно реже нарушения вызывают вирусные белки, напрямую ингибирующие синтез нуклеиновых кислот. Среди РНК-геномных вирусов наиболее быстрое и глубокое подавление синтезов нуклеиновых кислот вызывают пикорнавирусы, среди ДНК-геномных — покс- и герпесвирусы.
Во время репродукции вируса в клетке накапливаются вирусные компоненты, оказывающие токсическое и повреждающее действие на клеточные структуры. Например, цитотоксические свойства проявляют капсомеры некоторых аденовирусов, гликопротеины парамиксовирусов. В процессе вирусной инфекции также происходит повреждение мембран лизосом, содержимое которых высвобождается и осуществляет аутолиз клетки. Таким образом, гибель клеток наступает в результате сочетания раннего подавления синтеза клеточных компонентов, накопления токсических вирусных продуктов и повреждения лизосом.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
На уровне макроорганизма основные формы вирусных поражений принципиально не отличаются от таковых, наблюдаемых при инфицировании вирусами отдельных клеток.
Продуктивная вирусная инфекция с образованием дочерних популяций и характерными клиническими проявлениями возможна лишь при наличии в заражённом организме чувствительных клеток, в которых осуществляется репродуктивный цикл возбудителя. Например, возбудитель полиомиелита может реплицировать только в клетках ЖКТ и ЦНС приматов и человека.
Абортивная инфекция развивается при проникновении возбудителя в нечувствительные клетки (например, при попадании вируса лейкоза коров в организм человека) либо в клетки, не способные обеспечить полный репродуктивный цикл (например, находящиеся в стадии клеточного цикла G0). Способность клеток к поддержанию вирусспецифических репродуктивных процессов также подавляет ИФН, противовирусный эффект которого направлен против самых различных вирусов.

Персистирующая вирусная инфекция возникает при таком взаимодействии между вирусом и заражённой клеткой, когда в последней продолжается выполнение собственных клеточных функций. Если заражённые клетки делятся, образуется инфицированный клон. Таким образом, увеличение числа заражённых клеток способствует увеличению общей популяции возбудителя в организме. Тем не менее персистирующие вирусные инфекции обычно нарушают функции клеток, что в конце концов приводит к клиническим проявлениям. У человека развитие персисти-рующих инфекций в определённой степени зависит от возраста. Например, внутриутробное заражение вирусом коревой краснухи или цитомегаловирусом (ЦМВ) приводит к ограниченному по времени персистированию возбудителя. Появление симптоматики связано с возможностью плода развивать иммунные реакции на инфекционный агент.
Латентная (скрытая) вирусная инфекция. В то время как персистирующие инфекции сопровождаются постоянным высвобождением дочерних вирусных популяций, при латентных поражениях они образуются спорадически. Репродуктивный цикл подобных возбудителей резко замедляется на поздних стадиях и активируется под влиянием различных факторов. Латентные инфекции характерны для большинства герпесвирусов, вызывающих рецидивирующие и обычно не прогрессирующие заболевания.

Дремлющая (криптогенная) вирусная инфекция — форма проявления вирусной инфекции при которой возбудитель в неактивном состоянии находится в отдельных очагах (например, в нервных ганглиях). Клинически инфекция проявляется лишь при резком ослаблении защитных сил организма. Например, вирус герпеса 3 типа, вызывающий при первичном заражении ветряную оспу, пожизненно сохраняется в организме. Рецидив заболевания в форме опоясывающего лишая возможен лишь при нарушениях иммунного статуса (наиболее часто в пожилом возрасте).
Медленные вирусная инфекции характеризуются длительным инкубационным периодом (месяцы и годы), в течение которого возбудитель размножается, вызывая всё более явные повреждения тканей. Первоначально возбудитель размножается в ограниченной группе клеток, но постепенно инфицирует всё большее их число. Заболевания заканчиваются развитием тяжёлых поражений и смертью больного. К медленным вирусным инфекциям относят подострый склерозирующий панэнцефалит, ВИЧ-инфекцию и др.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Взаимодействие вируса с клеткой хозяина - это сложный «ногоступенчатый процесс, который начинается с адсорбции вирусных частиц на рецепторах клетки хозяина и продолжается после их проникновения внутрь клетки. В результате такого взаимодействия мовивается либо продуктивная, либо абортивная, либо интегративная форма клеточной инфекции. При продуктивной форме происходит размножение, точнее, репродукция (лат. reproduce - воспроизводить) вируса, при абортивной - ее нарушение на одном из этапов, при интегративной - интеграция вирусной нуклеиновой кислоты в клеточный геном.
Продуктивная инфекция. Репродукция вирусов
Как отмечалось выше, вирусы являются самореплицирующейся формой, неспособной к бинарному делению, в отличие от микроорганизмов с клеточной организацией. В 50-х годах было установлено, что размножение, или репродукция, вирусов происходит путем репликации их нуклеиновой кислоты и биосинтеза белков с последующей самосборкой вириона. Этот процесс происходит в разных частях клетки - ядре или цитоплазме, вследствие чего получил название дизъюнктивного, т.е. разобщенного размножения.
Вирусная репродукция представляет собой уникальную форму выражения чужеродной (вирусной) информации в клетках человека и животных, насекомых, растений и бактерий, которая состоит в подчинении клеточных матрично-генетических механизмов вирусной информации.
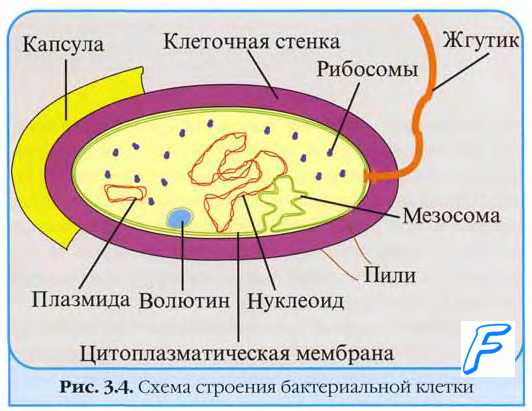
1-я стадия - адсорбция - характеризуется прикреплением вириона к клеточным рецепторам, представляющим собой гликопротеины клеточной мембраны, содержащей нейраминовую кислоту. Такие рецепторы имеются у ряда клеток, в частности эритроцитов, на которых адсорбируются многие вирусы. Для орто- и парамиксовирусов специфическими рецепторами являются гликолипиды, содержащие сиаловую кислоту (ганглиозиды), для других - белки или липиды клеточной мембраны. Рецепторами вирусов являются так называемые прикрепительные белки, располагающиеся в составе капсидов простых вирионов и суперкапсидов сложных вирионов. Они могут иметь форму нитей (фибры у аденовирусов) или шипов (гликопротеиновые образования на внешней оболочке орто- и парамиксо-, рабдо-, арено- и буньявирусов). Первый этап адсорбции определяется неспецифическими силами межмолекулярного притяжения, второй - специфической структурной гомологией или комплементарностью рецепторов чувствительных клеток и вирусов.
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
2-я стадия - проникновение вируса в клетку хозяина - происходит несколькими путями.
Рецепторно-опосредованный эндоцитоз характеризуется образованием в месте взаимодействия вириона с клеточным рецептором окаймленных пузырьков, в формировании которых принимают участие белки-клатрины.
Виропексис. Этим путем в клетку проникают сложноустроен-ные вирусы. Он заключается в слиянии мембран - вирусного суперкапсида с клеточной или ядерной мембраной. Данный процесс происходит при помощи специального бека слияния - F-белка, который находится в суперкапсиде. В результате виропексиса капсид оказывается в клетке хозяина, а суперкапсид вместе с белком встраивается в ее плазматическую мембрану (вследствие чего клетка приобретает способность сливаться с другими клетками, что приводит к передаче вируса этим клеткам).
Фагоцитоз. Данным путем вирус проникает в фагоцитирующие клетки, что приводит к незавершенному фагоцитозу.
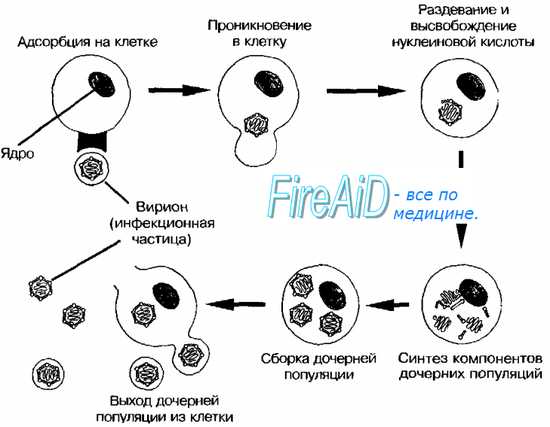
3-я стадия - транспорт вируса внутри клетки. Он происходит с помощью внутриклеточных мембранных пузырьков, в которых вирус переносится на рибосомы, эндоплазматическую сеть или в ядро.
5-я стадия называется эклипс-фазой, которая характеризуется исчезновением вириона, поскольку он перестает обнаруживаться при элданой микроскопии. В эту стадию начинается синтез компонентов вириона, т.е. его репродукция. Она носит дизъюнктивный (раздельный) характер, поскольку компоненты вириона синтезируются в разных частях клетки: белки на рибосомах, нуклеиновые кислоты в ядре или цитоплазме. Вирус использует для этого генетический аппарат клетки, подавляя необходимые ей самой синтетические реакции.
Эта стадия начинается с транскрипции и репликации вирусного генома. Транскрипция вирусного генома двунитевых ДНК-содержащих вирусов происходит, так же как и клеточного генома, по триаде ДНК- иРНК- белок. Различия касаются только происхождения фермента ДНК-зависимой РНК-полимеразы, необходимой для данного процесса. У вирусов, геном которых транскрибируется в цитоплазме клетки хозяина (например, вирус оспы), имеется собственная вирусспецифическая РНК-полимераза. Вирусы, геномы которых транскрибируются в ядре (папова- и аденовирусы, вирусы герпеса), используют содержащуюся там клеточную РНК-полимеразу II или III.
Теневая фаза ( фаза эклипса )
После депротеинизации вирусы невозможно выделить из культуры клеток. Этот этап репродукции известен как теневая фаза, или фаза эклипса [от англ. eclipse, затмение].
Она включает репликацию нуклеиновых кислот вируса и синтез вирусных белков. Теневая фаза не происходит при температуре 0-4 °С (исключая вирус гриппа). Различия в энергетических потребностях для теневой фазы разных групп вирусов указывают на возможное участие в этом процессе различных клеточных реакций. Теневая фаза заканчивается после образования составных компонентов вируса, необходимых для сборки дочерних популяций.
ЦПД — видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения от стекла), возникающие в результате внутриклеточной репродукции вирусов.
Образование вирусных частиц
Образование дочерних вирусных частиц в заражённой клетке подразумевает необходимость трёх процессов:
экспрессия генетического материала в виде его транскрипции и последующей трансляции, что приводит к появлению вирусных белков;
синтез генетического материала вируса (репликация);
сборка из генетического материала и вирусных белков дочерних популяций.
Следует помнить, что генетическим материалом вирионов может быть либо ДНК, либо РНК
У РНК-содержащих вирусов транскрипция их генома осуществляется несколькими путями:
1. Вирусы с негативным геномом (минус-нитевые), к которым относятся орто-, парамиксо- и рабдовирусы, имеют в своем составе вирусспецифическую РНК-полимеразу или транскриптазу. Они синтезируют иРНК на матрице геномной РНК. Подобный фермент отсутствует в нормальных клетках, но синтезируется клетками, зараженными вирусами. Он находится в составе как однонитевых, так и двунитевых РНК-содержащих вирусов.
2. У вирусов с положительным геномом (плюс-нитевые), к которым относятся пикорна-, тогавирусы и др., функцию иРНК выполняет сам геном, который транслирует содержащуюся в нем информацию на рибосомы клетки хозяина.
Транскрипция -РНК-содержащих вирусов и вирусов, имеющие две нити РНК. Функции мРНК выполняют транскрипты, комплементарные -РНК вириона. Поэтому у таких вирусов транскрипция существует как самостоятельный этап репродуктивного цикла. Для образования транскриптов в составе вирионов имеется собственная РНК-полимераза (транскриптаза).
Транскрипция +РНК-содержащих вирусов. Функции мРНК выполняет геном (+РНК), поэтому у таких вирусов для синтеза вирусных белков (трансляция) нет необходимости в процессе транскрипции. Другими словами, у +РНК-содержащих вирусов транскрипция отсутствует.
Транскрипция ДНК-вирусов. Транскрипция — самостоятельный этап репродуктивного цикла, так как геном ДНК-вирусов должен транскрибироваться для образования мРНК. Вирусы, репродуцирующиеся в ядре (например, герпес- и аденовирусы) для этой цели используют клеточную ДНК-зависимую РНК-полимеразу (транскриптазу). Вирусы, репродуцирующиеся в цитоплазме (например, поксвирусы) лишены такой способности и содержат (как и вирусы с -РНК) собственную транскриптазу.
Читайте также: