
Основные схемы репликации вирусов при продуктивной инфекции
Обновлено: 26.04.2024
Репликация вирусов. Как размножаются вирусы?
Вирионы представляют собой инертные формы, которые сами по себе не размножаются, а лишь существуют для передачи от одного организма другому и для перехода из одной клетки хозяина в другую и сохранения во внешней среде. В последнем случае они как бы напоминают роль споровых форм микроорганизмов.
Одни вирусы имеют широкий спектр хозяев, другие заражают лишь клетки одного типа определенных видов животных. Способность клеток и организма заражаться вирусом называют, соответственно, чувствительностью и восприимчивостью. Когда восприимчивый организм вступает в контакт с вирусом, первоначально заражаются лишь чувствительные клетки у входных ворот инфекции (например, миндалины). Однако для клинического проявления инфекции этого может оказаться недостаточно, так как болезнь возникает вследствие заражения клеток-мишеней (например, нейронов, гепатоцитов) после того, как вирус размножился на месте внедрения в организм. В ряде случаев клетки-мишени располагаются у входных ворот инфекции (например, кишечные, респираторные инфекции).
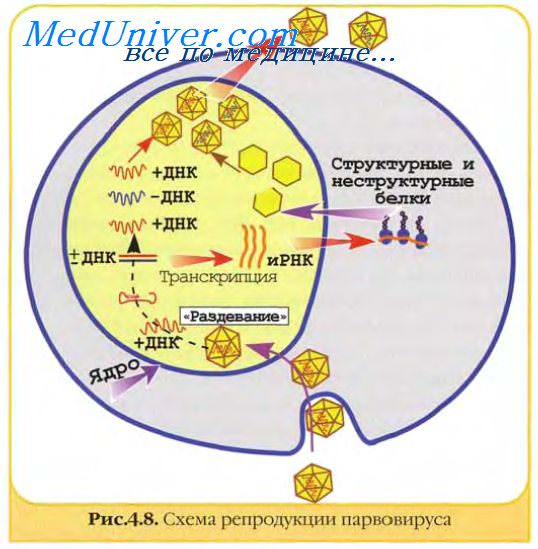
События, происходящие в инфицированной клетке, могут существенно различаться и конечный результат их также может быть различным — от размножения вируса (продуктивная инфекция) с разрушением или/без разрушения клеток до трансформации клеток, которые приобретают способность к неограниченному делению.
Многие цитопатогенные вирусы обладают способностью к установлению персистентной инфекции в чувствительных клетках. Молекулярные механизмы персистенции вирусов в клеточных культурах окончательно не выяснены.
Репликация вирусов занимает центральное место в молекулярной биологии. Основные закономерности репликации вирусов первоначально были изучены в опытах с бактериофагами, а с появлением культур клеток млекопитающих эти методы были адаптированы к вирусам животных.
В начале инфекций в клетку проникает вирусный геном (РН К или ДНК), часто вместе с необходимыми вирионными белками. Несмотря на существенные различия в стратегии размножения разных семейств ДНК- и РНК-вирусов, особенно в отношении транскрипции и репликации их геномов, этот процесс имеет общие характерные особенности, свойственные всем вирусам. Главная из них состоит в том, что способность вирусов к размножению и судьба зараженных клеток зависят от синтеза и функции продуктов вирусных генов - белков, которые обеспечивают: а) репликацию вирусного генома и синтез вирусных белков, б) упаковку генома в вирусные частицы (вирионы) и в) изменение структуры и (или) функции зараженных клеток.
Стратегия, применяемая вирусами для обеспечения этих функций, варьирует. Иногда вирусные белки (папиллома и полиомавирусы) просто помогают ферментам клетки хозяина реплицировать вирусный геном. В большинстве случаев вирусные белки сами обеспечивают репликацию вирусного генома, но даже наиболее независимые вирусы , как правило, используют при этом по крайней мере несколько белков хозяина.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
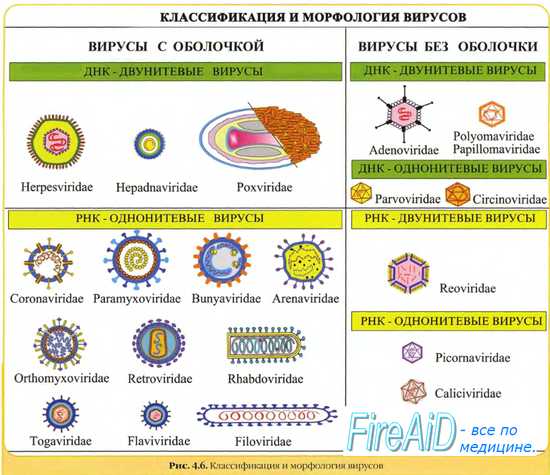
В зависимости от типа генетического материала (ДНК или РНК), образование дочерних копий геномов протекает по-разному.
У ДНК-геномных вирусов репликация вирусных ДНК принципиально сходна с репликацией клеточных ДНК.
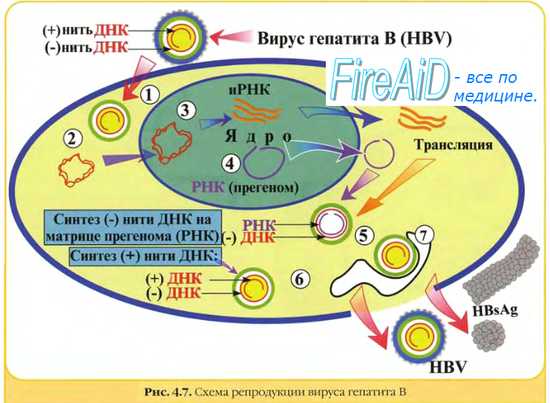
Репликацию РНК-геномных вирусов осуществляют вирусные РНК-зависимые РНК-полимеразы (репликазы). Исключение составляют ретровирусы, их +РНК служит матрицей для синтеза ДНК Синтез ДНК на матрице РНК осуществляет вирусная РНК-зависимая ДНК-полимераза (обратная транскриптаза), необходимая для переписывания информации с РНК на ДНК. Синтезируемая вирусная ДНК интегрируется в клеточный геном в форме ДНК-провируса.
Репликация однонитевых РНК вирусов. Репликация протекает в два этапа: первый включает образование матрицы, комплементарной геному; второй — образование копий РНК с этой матрицы. При репликации +РНК-вирусов количество копий -РНК (на матрице родительской нити +РНК) строго контролируется, а количество копий +РНК (с матрицы синтезированной нити -РНК) не контролируется.
Репликация двухнитевых РНК вирусов. В качестве матрицы для синтеза +РНК вирусные репликазы используют минус-нить РНК и наоборот. Часть молекул -РНК соединяется с +РНК и образует двухнитевую молекулу РНК, а другая часть молекул -РНК функционирует как матрица для синтеза мРНК.
Сборка вирусов
У просто устроенных вирусов, состоящих из нуклеиновой кислоты и нескольких белков, сборка состоит из упорядоченного взаимодействия этих молекул. У сложно устроенных вирусов сборка дочерних популяций протекает многоступенчато.
Взаимодействие нуклеиновых кислот с внутренними и оболочечными белками приводит к образованию нуклеокапсидов, или сердцевин. В процессе образования «одетых - вирусов полные нуклеокапсиды упорядочение выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками (М-белками). При нарушениях процесса самосборки могут образовываться пустые капсиды либо комплексы нуклеиновых кислот с внутренними белками.
Высвобождение дочерних вирионов из клетки
Высвобождение дочерних вирионов — конечная стадия репродуктивного цикла. Вирусы, лишённые суперкапсида, и поксвирусы обычно высвобождаются быстро; выход дочерних популяций сопровождается разрушением цитоплазматической мембраны (ЦПМ) и лизисом клетки. Вирусы, содержащие суперкапсид, высвобождаются медленнее. Модифицированные участки мембраны с заключёнными в них вирионами выпячиваются наружу и затем отпочковываются. Принцип высвобождения дочерних популяций почкованием во многом сходен с процессами, направленными на отторжение непригодного для клетки материала или обновление клеточных мембран. При высвобождении почкованием изменённая клетка иногда может сохранять жизнеспособность.
- Вернуться в оглавление раздела "Микробиология."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Репликация (вирусология) – это процесс самовоспроизведения нуклеиновых кислот, генов, хромосом. В его основе лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [2] .
Механизм репликации, также, как и механизм транскрипции, зависит от типа геномной НК вирусов [1] .
Механизм репликации ДНК-вирусов
При репликации двухнитчатых ДНК-вирусов матрицей для синтеза комплементарных дочерних цепей служит каждая из цепей родительской ДНК. Для репликации геномных ДНК большинство вирусов используют клеточные ферменты, в частности ДНК-зависимые ДНК-полимеразы. Некоторые крупные вирусы кодируют собственные репликазы [3] [1] .
Вирусы, содержащие плюс-ДНК, начинают репликацию с синтеза комплиментарной минус-ДНК нити. Образуется двухспиральная репликативная форма, которая выступает в роли матрицы для синтеза новых плюс-цепей. Последние, вытесняются из репликативной формы и заключаются в белковую оболочку, превращаясь в вирионы [1] .
Принципиальное отличие в репликации однонитевых ДНК содержащих вирусов от двухнитевых состоит в том, что в первом случае для формирования дочерних молекул используется только минус-цепь [3] [1] .
Механизм репликации однонитевых РНК-вирусов
Репликация однонитевых РНК-вирусов в общих чертах сходна с репликацией однонитевых ДНК-вирусов. В начале процесса однонитевая родительская плюс-РНК служит матрицей для синтеза дочерней минус-РНК. В результате формируется двухспиральная репликативная форма [1] .
Но за долго до окончания синтеза первой дочерней РНК, на репликативной форме инициируется синтез второй и последующих дочерних плюс-нитей. Этот процесс приводит к преобразованию репликативной формы в новую форму, именуемую репликативным предшественником [1] .
Механизм репликации двухцепочечных РНК-вирусов
РНК-вирусы с двухцепочечным геномом используют в качестве матриц для репликации только минус-нити. Эти вирусы отличает то, что уже на ранних стадиях инфекции в клетках-хозяевах накапливаются плюс-РНК. Некоторые из них становятся матрицами для репликации. Другие используются как иРНК [3] [1] .
Репликация ретровирусов
Репликация ретровирусов (семейство вирусов для которых характерен процесс обратной транскрипции) наблюдается на матрице провирусов (генома вируса, встроенного в ДНК клетки хозяина) после их индукции (процесса исключения провируса из состава хромосомы клетки-хозяина) [1] .
Репликация провирусов происходит с помощью клеточных ферментов. В результате синтезируются многочисленные молекулы плюс-РНК и необходимые для сборки вирионов белки и ферменты [1] .
Репродукция вируса – это процесс размножения вирусных частиц в чувствительных к ним клетках. Репродуцируются только вирулентные вирусы, обладающие высокой степенью патогенности [3] .
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций [3] .
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина [2] .
- Все вирусы, содержащие молекулу ДНК, кроме вирусаоспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирусоспы размножается только в цитоплазме клетки [2] .
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами [2] .
- Белки вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению) [2] .
- Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование (присоединении к полипептиду углеводных остатков) [2] .

Этапы репродукции (жизненного цикла) вируса

1. Адсорбция вируса на мембране клетки.; 2. Проникновение вируса в клетку.; 3. Депротеинизация.; 4. Синтез компонентов вирусов.; 5. Формирование дочерних вирионов.; 6. Выход вирионов [2] .
Этапы репродукции вируса
Репродукцию или жизненный цикл вируса делят на шесть последовательных этапов:
- Адсорбция на мембране клетки [2] .
- Проникновение в клетку [2] .
- Депротеинизация [2] .
- Синтез компонентов вирусов[2] .
- Формирование дочерних вирионов[2] .
- Выход вирионов[2] .

Процесс адсорбции вириона на поверхности клетки

1. Вирион.; 2. Клеточный рецептор.; 3. Прикрепительный белок; 4. Клеточная мембрана [2] .
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона [2] .
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур [2] .
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно [2] .
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 [2] .

Проникновение вируса в клетку (Путь I)

Слияние вирусной оболочки с клеточной мембраной
1. Вирион.; 2. Инфицируемая клетка.; 3. Ядро [2] .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона [2] . Существует два пути:
- Путь I – слиянии вирусной оболочки с клеточной мембраной [2] .
- Путь II – рецептор-опосредованный эндоцитоз [2] .
Путем слияния суперкапсида с клеточной мембраной (путь I) в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки [2] .
Путем рецептор-опосредованного эндоцитоза (путь II) в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении, вирусный геном в цитоплазме клетки освобождается из эндосомы [2] .

Проникновение вируса в клетку (Путь II)

1. Вирион.; 2. Клетка.; 3. Эндосома.; 4. Ядро [2] .
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы) [2] .
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона [2] .
Синтез компонентов вируса
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [1] .
Место синтеза компонентов дочерних вирионов зависит от типа генома:
- реализация генетической информации у ДНК-содержащих вирусов идет по пути: ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом идет по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок;
- РНК-содержащие ретровирусы идут по следующему пути передачи информации: РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок [2] .
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона [2] .
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках [2] .
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК [2] .
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается [2] .
РНК-содержащие ретровирусы отличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза [2] [4] .
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков, и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры [2] .
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более [2] .
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования[2] .
Путь лизиса клетки тесно связан ее деструкцией. Он характерен для безоболочечных вирусов, не имеющих суперкапсидной оболочки (суперкапсида) [2] .
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы, высвобождаются в течении 2–6 часов. В начале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида [2] .
Читайте также: