
Синтез днк на матрице и-рнк у вирусов
Обновлено: 12.05.2024
ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
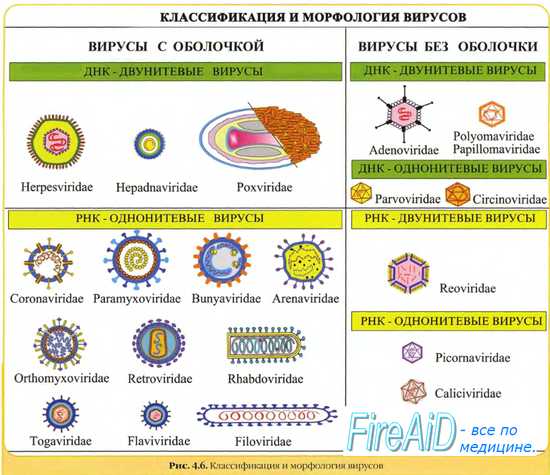
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
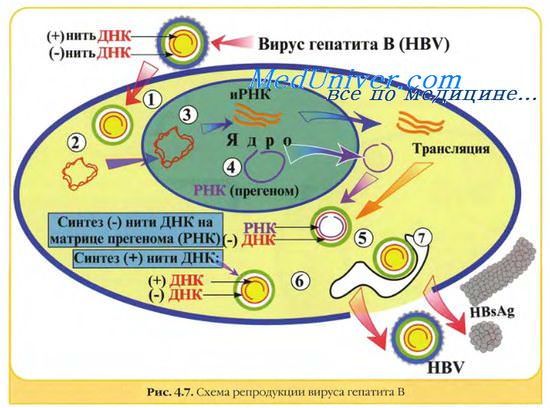
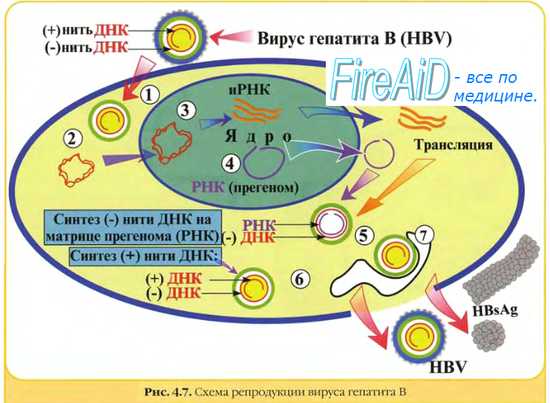
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
В зависимости от типа генетического материала (ДНК или РНК), образование дочерних копий геномов протекает по-разному.
У ДНК-геномных вирусов репликация вирусных ДНК принципиально сходна с репликацией клеточных ДНК.
Репликацию РНК-геномных вирусов осуществляют вирусные РНК-зависимые РНК-полимеразы (репликазы). Исключение составляют ретровирусы, их +РНК служит матрицей для синтеза ДНК Синтез ДНК на матрице РНК осуществляет вирусная РНК-зависимая ДНК-полимераза (обратная транскриптаза), необходимая для переписывания информации с РНК на ДНК. Синтезируемая вирусная ДНК интегрируется в клеточный геном в форме ДНК-провируса.
Репликация однонитевых РНК вирусов. Репликация протекает в два этапа: первый включает образование матрицы, комплементарной геному; второй — образование копий РНК с этой матрицы. При репликации +РНК-вирусов количество копий -РНК (на матрице родительской нити +РНК) строго контролируется, а количество копий +РНК (с матрицы синтезированной нити -РНК) не контролируется.
Репликация двухнитевых РНК вирусов. В качестве матрицы для синтеза +РНК вирусные репликазы используют минус-нить РНК и наоборот. Часть молекул -РНК соединяется с +РНК и образует двухнитевую молекулу РНК, а другая часть молекул -РНК функционирует как матрица для синтеза мРНК.
Сборка вирусов
У просто устроенных вирусов, состоящих из нуклеиновой кислоты и нескольких белков, сборка состоит из упорядоченного взаимодействия этих молекул. У сложно устроенных вирусов сборка дочерних популяций протекает многоступенчато.
Взаимодействие нуклеиновых кислот с внутренними и оболочечными белками приводит к образованию нуклеокапсидов, или сердцевин. В процессе образования «одетых - вирусов полные нуклеокапсиды упорядочение выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками (М-белками). При нарушениях процесса самосборки могут образовываться пустые капсиды либо комплексы нуклеиновых кислот с внутренними белками.
Высвобождение дочерних вирионов из клетки
Высвобождение дочерних вирионов — конечная стадия репродуктивного цикла. Вирусы, лишённые суперкапсида, и поксвирусы обычно высвобождаются быстро; выход дочерних популяций сопровождается разрушением цитоплазматической мембраны (ЦПМ) и лизисом клетки. Вирусы, содержащие суперкапсид, высвобождаются медленнее. Модифицированные участки мембраны с заключёнными в них вирионами выпячиваются наружу и затем отпочковываются. Принцип высвобождения дочерних популяций почкованием во многом сходен с процессами, направленными на отторжение непригодного для клетки материала или обновление клеточных мембран. При высвобождении почкованием изменённая клетка иногда может сохранять жизнеспособность.
- Вернуться в оглавление раздела "Микробиология."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Репликация вирусных РНК молекул. Этапы репликации РНК генома вирусов.
Репликация вирусных РНК является уникальным феноменом. Существенное отличие механизма синтеза вирусных РНК от механизма синтеза клеточных РНК состоит в том, что в качестве матрицы в первом случае используется РНК, а во втором — ДНК.
Для транскрипции РНК на РНК-матрице необходима вирионная РНК-зависимая РНК-полимераза. Репликация вирусной РНК требует, прежде всего, синтеза комплементарной РНК, которая затем служит матрицей для производства большого количества вирусной РНК.
Когда вирусная РНК имеет отрицательную полярность (орто-, парамиксо-, рабдо-, фило-, борна-, арена- и буньявирусы), комплементарная РНК будет иметь положительную полярность и РНК-полимераза, подобно вирионной транскриптазе, используется для первичной транскрипции мРНК.
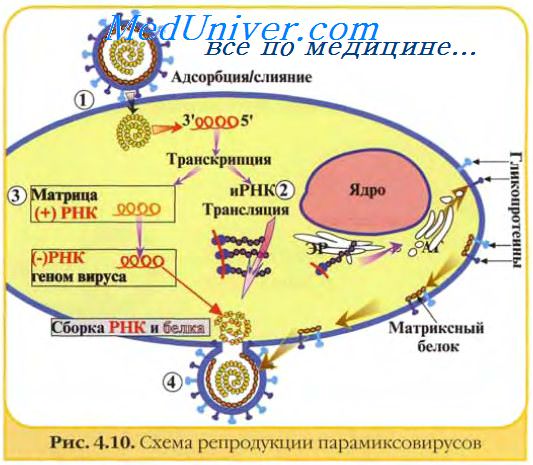
Так как большинство транскриптов, синтезируемых на каждой вирусной (-)цепи РНК, являются молекулами субгеномной РНК, некоторые полноразмерные цепи служат матрицами для синтеза (репликации) вирусной РНК. Некоторые вирусы для транскрипции и репликации используют различные РНК-полимеразы, тогда как в других случаях одни и те же ферменты могут выполнять различные функции.
У многих РНК-вирусов, (пикорна-, калици-, астро-, тога-, флави-, корона-, артери-, нодавирусы) комплементарная РНК является отрицательно полярной. На одной комплементарной РНК-матрице может транскрибироваться одновременно несколько молекул вирусной РНК, а на каждом РНК-транскрипте начинается продукция полимеразы. Образуется структура, известная как реплика-тивный посредник, — частично двуцепочечная структура с одноцепочечными хвостами.
Для начала репликации РНК пикорнавирусов и калицивирусов, а также ДНК аденовирусов требуется небольшой белок, связанный ковалентно с 5'-концом вновь синтезированных (+) или (—) цепей РНК, так же как с родительской вирионной РНК, но не с мРНК.
Вновь синтезированные (+)РНК могут иметь разное назначение: включаться в репликативный комплекс и служить матрицей для синтеза комплементра-ных (—)РНК; выполнять функции мРНК; включаться в качестве генома в новые вирионы. Механизм, определяющий судьбу вновь синтезированных (+)РНК, не известен.
Ретровирусы имеют геномную (+) одноцепочечную РНК. В отличие от других РНК-вирусов, они реплицируются посредством ДНК-посредника. Вирионная обратная транскриптаза, используя РНК-молекулу как праймер, создает односпиральную ДНК-копию. Затем, функционируя как рибонуклеаза, тот же самый фермент удаляет родительскую молекулу РНК из ДНК-РНК-гибрида и копирует одноцепочечную ДНК-цепь, чтобы образовать линейную двуцепочечную ДНК, которая содержит дополнительную последовательность, известную как длинный концевой повтор (LTR) на каждом конце.
Эта двуцепочечная ДНК затем циркулирует и интегрирует с клеточной хромосомальной ДНК. Вирусная РНК транскрибируется с интегрированной (провирусной) ДНК.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Репликация вирусных РНК молекул. Этапы репликации РНК генома вирусов.
Репликация вирусных РНК является уникальным феноменом. Существенное отличие механизма синтеза вирусных РНК от механизма синтеза клеточных РНК состоит в том, что в качестве матрицы в первом случае используется РНК, а во втором — ДНК.
Для транскрипции РНК на РНК-матрице необходима вирионная РНК-зависимая РНК-полимераза. Репликация вирусной РНК требует, прежде всего, синтеза комплементарной РНК, которая затем служит матрицей для производства большого количества вирусной РНК.
Когда вирусная РНК имеет отрицательную полярность (орто-, парамиксо-, рабдо-, фило-, борна-, арена- и буньявирусы), комплементарная РНК будет иметь положительную полярность и РНК-полимераза, подобно вирионной транскриптазе, используется для первичной транскрипции мРНК.
Так как большинство транскриптов, синтезируемых на каждой вирусной (-)цепи РНК, являются молекулами субгеномной РНК, некоторые полноразмерные цепи служат матрицами для синтеза (репликации) вирусной РНК. Некоторые вирусы для транскрипции и репликации используют различные РНК-полимеразы, тогда как в других случаях одни и те же ферменты могут выполнять различные функции.
У многих РНК-вирусов, (пикорна-, калици-, астро-, тога-, флави-, корона-, артери-, нодавирусы) комплементарная РНК является отрицательно полярной. На одной комплементарной РНК-матрице может транскрибироваться одновременно несколько молекул вирусной РНК, а на каждом РНК-транскрипте начинается продукция полимеразы. Образуется структура, известная как реплика-тивный посредник, — частично двуцепочечная структура с одноцепочечными хвостами.
Для начала репликации РНК пикорнавирусов и калицивирусов, а также ДНК аденовирусов требуется небольшой белок, связанный ковалентно с 5'-концом вновь синтезированных (+) или (—) цепей РНК, так же как с родительской вирионной РНК, но не с мРНК.
Вновь синтезированные (+)РНК могут иметь разное назначение: включаться в репликативный комплекс и служить матрицей для синтеза комплементра-ных (—)РНК; выполнять функции мРНК; включаться в качестве генома в новые вирионы. Механизм, определяющий судьбу вновь синтезированных (+)РНК, не известен.
Ретровирусы имеют геномную (+) одноцепочечную РНК. В отличие от других РНК-вирусов, они реплицируются посредством ДНК-посредника. Вирионная обратная транскриптаза, используя РНК-молекулу как праймер, создает односпиральную ДНК-копию. Затем, функционируя как рибонуклеаза, тот же самый фермент удаляет родительскую молекулу РНК из ДНК-РНК-гибрида и копирует одноцепочечную ДНК-цепь, чтобы образовать линейную двуцепочечную ДНК, которая содержит дополнительную последовательность, известную как длинный концевой повтор (LTR) на каждом конце.
Эта двуцепочечная ДНК затем циркулирует и интегрирует с клеточной хромосомальной ДНК. Вирусная РНК транскрибируется с интегрированной (провирусной) ДНК.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: