
Вирусная рнк зависимая рнк полимераза
Обновлено: 25.04.2024
РНК-вирусы. Особенности размножения РНК-вирусов.
Пикорна-, калици-, астро-, тога- и флавивирусы реплицируются наиболее коротким путем: их (+)РНК геном функционирует непосредственно как мРНК.
Геномы пикорна- и флавивирусов функционируют как единая полицистронная мРНК, транслирующаяся прямо в единый полипротеин, который впоследствии расщепляется с образованием индивидуальных структурных и неструктурных белков. Одним из них является РНК-зависимая РНК-полимераза, которая реплицирует вирусный геном. Вирусная (+)РНК транскрибируется в комплиментарную (—)РНК-копию, которая служит матрицей для синтеза новых цепей (+)РНК (рис. 6), или может использоваться в качестве мРНК-матриц для синтеза новых минус-цепей и геномной РНК вирусного потомства.
Тога-, корона- и калицивирусы отличаются от пикорнавирусов тем, что на начальном этапе инфекции экспрессируется лишь часть геномной РНК с образованием белков. Последние осуществляют синтез минус-цепи, являющейся матрицей для синтеза различных по размеру классов молекул плюс-РНК. Полипротеины, образующиеся на коротких молекулах мРНК, расщепляются на структурные вирионные белки. Полноразмерные плюс-РНК упаковываются в вирионы.
У тогавирусов транслируется только около 2/3 вирусной РНК (5' -конец); образующийся полипротеин расщепляется на неструктурные белки, которые необходимы для транскрипции и репликации РНК. Вирусная РНК-полимераза синтезирует полноразмерную (—)РНК, на которой затем синтезируются два вида (+)РНК: полноразмерная вирионная РНК, предназначенная для включения в вирионы, и РНК, длина которой равна 1/3, и которая является колинеарной с 3'-концом вирусной РНК и транслируется в полипротеин, который расщепляется на структурные белки. У калицивирусов образуются полигеномные и субгеномные мРНК.
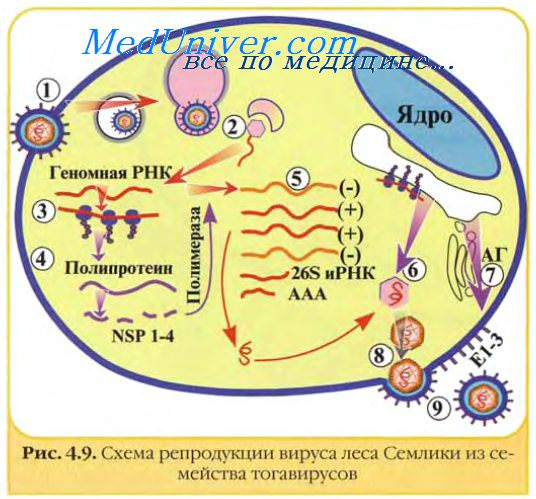
Корона- и артеривирусы демонстрируют необычную стратегию транскрипции: первоначально часть вирионной (+) РНК функционирует как мРНК и транслируется с образованием РНК-полимеразы, которая затем синтезирует полногеномную (—)РНК. На этой (—)РНК транскрибируется гнездо субгеномных мРНК с общими 3'-концами. Транслируются только 5'-концевые последовательности каждого члена этого гнезда транскриптов.
Главным отличием вирусов с позитивным геномом является их способность синтезировать ферменты, ответственные за репликацию вирусного генома. Поэтому РНК, выделенная из таких вирусов, инфекционна. Второе отличие состоит в монолитности вирусного генома. Поэтому первичный продукт трансляции обеих РНК (геномной и мРНК) представляет собой единый белок, который в дальнейшем расщепляется на индивидуальные вирусные белки (в том числе структурные).
Ортомиксо-, бунья- и аренавирусы. Геном представлен (—)РНК. Каждый геномный сегмент транскрибируется отдельно вирионной РНК-транскриптазой и транслируется в один или несколько белков. Эти вирусы характеризуются тем, что их геномная РНК выполняет две матричные функции: в процессе транскрипции и репликации. Транскрипция вирусного генома — первое событие после проникновения вируса в клетку, в результате которого образуются моноцистронные мРНК, кодирующие один белок. Репликацию начинают вновь синтезированные вирусные белки, приводящие к образованию плюс-цепи, которая служит матрицей для синтеза геномной (-)РНК.
Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от (+)РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК.
Следует отметить, что S-сегмент РНК аренавирусов и некоторых буньявирусов является двуполярным, т.е. одна часть сегмента имеет (+) полярность, другая (—) полярность. Стратегия репликации двуполярных РНК-вирусов соответствует полярности их геномов и является смешанной, присущей стратегии репликации (+)РНК и (-)РНК вирусов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Обзор
здесь и далее рисунки Андрея Занкевича
Автор
Редактор

Генеральный партнер конкурса — ежегодная биотехнологическая конференция BiotechClub, организованная международной инновационной биотехнологической компанией BIOCAD.

Спонсор конкурса — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Напомним, что РНК (рибонуклеиновая кислота) — это такая молекула, представляющая собой одну цепочку нуклеотидов. В составе каждого нуклеотида присутствует остаток моносахарида рибозы. На сегодняшний день известно множество разных типов РНК, которые выполняют совершенно разные функции: от кодирования клеточных белков (мРНК) до противовирусной защиты (некоторые микроРНК) [1–5]. РНК, входящие в состав многих вирусов, могут иметь ряд оригинальных функций, таких как регуляция времени экспрессии различных вирусных генов путем изменения пространственной организации цепи РНК или привлечение клеточных белковых комплексов.
Но бывает ли такое, что в пределах одной молекулы РНК одна ее часть, кодирующая некоторый белковый продукт, имеет положительную полярность, в то время как другая часть цепи представлена участком отрицательной полярности, кодирующим другой белок? Могла ли такая молекула возникнуть в процессе эволюции живых форм?
Ответ — да! И для того, чтобы разобраться, как функционируют такие молекулы, нам предстоит погрузиться в таинственный мир вирусов.
Давным-давно, в далекой-далекой галактике.
Как известно, все формы жизни обладают определенной наследственностью, которая определяет степень генетической идентичности живых объектов. В качестве молекул, ответственных за поддержание такой наследственной идентичности, выступают нуклеиновые кислоты (ДНК и РНК). Клеточные формы жизни для хранения и передачи информации используют только один тип нуклеиновых кислот — дезоксирибонуклеиновую кислоту (ДНК), в то время как вирусы в качестве основной молекулы наследственности могут использовать либо ДНК, либо РНК.
В случае ДНК-содержащих вирусов реализуется, как правило, Центральная догма молекулярной биологии в классическом виде: попав в клетку, ДНК удваивается в процессе репликации вирусного генома, на матрице ДНК в ходе транскрипции синтезируются мРНК, которые затем прочитываются рибосомой, синтезирующей по ним вирусные белки, то есть осуществляется трансляция. Далее вирусные белки ассоциируются с ДНК-геномом вируса в вирусную частицу (вирион), которая способна заражать новые клетки.
Очевидно, что РНК-содержащие вирусы используют иные стратегии размножения и, следовательно, реализации своего генома. Непривычные для большинства биологов молекулярные механизмы, которые используются такими вирусами, вероятно, унаследованы от далеких предков из того самого РНК-мира.
Размножение РНК-вирусов подразумевает использование разных типов РНК:
- геномная РНК находится внутри вириона, в зависимости от конкретного вируса, она может быть представлена (+)РНК, (–)РНК, (±)РНК, либо двухцепочечной РНК;
- комплементарная геномной РНК антигеномная РНК образуется в процессе репликации вирусов с одноцепочечным РНК-геномом и обладает полярностью, противоположной геномной РНК;
- субгеномная РНК (вирусная мРНК) имеет (+)полярность и является продуктом транскрипции геномной или антигеномной РНК. Как и подобает мРНК, субгеномная РНК участвует в процессе трансляционного синтеза белка.
Немного истории
Первым найденным РНК-вирусом стал бактериофаг f2, инфицирующий бактерию кишечную палочку (Escherichia coli) [7]. Выделенная геномная РНК фага f2 имела свойства мРНК, то есть она распознавалась рибосомой и могла транслироваться. На родственном РНК-бактериофаге Qβ была изучена РНК-зависимая РНК-полимераза (RdRp), которая, как оказалось, может синтезировать цепь РНК на матрице другой цепи РНК, то есть осуществлять репликацию вирусного РНК-генома! В ходе работы с РНК-бактериофагами f2 и его родственником Qβ были получены общие представления о биологии таких РНК-вирусов [8].
После РНК-бактериофагов были найдены (+)РНК-вирусы животных, такие, как вирус полиомиелита [9], [10], представитель группы пикорнавирусов. Подобные вирусы не содержат репликативных белков в составе вирусной частицы (вириона).
Встречаются вирусы, геном которых представлен двухцепочечной РНК. Как и в случае (–)РНК-вирусов, у дцРНК-вирусов во время репликации в клетке первым делом происходит синтез (+)цепи. Вирусные частицы этой группы также включают в свой состав RdRp.
Вирусы с двусмысленным РНК-геномом из семейства Bunyaviridae

Рисунок 1. Схематичное изображение структуры вириона флебовирусов
В семействе Bunyaviridae роды Phlebovirus, Tospovirus и Tenuivirus являются вирусами с двусмысленным РНК-геномом и, в отличие от остальных представителей семейства, имеют чуть более длинный S-сегмент генома (РНК S) (±)полярности. Род Tospovirus имеет вдобавок (+)участок на РНК M, который делает и эту РНК амбисенсной.
Флебовирусы
Вирусы рода Phlebovirus выделяют практически по всему миру и относят к нетаксономической группе арбовирусов, распространяющихся в членистоногих переносчиках и в позвоночных, на которых питаются переносчики. Члены этого рода переносятся кровососущими членистоногими. Инфекции не обходят стороной человека: вирусы сицилийской и неаполитанской москитных лихорадок широко распространены по территории Средиземноморья [15]. Среди симптомов таких инфекций — продолжительная сильная лихорадка, тошнота, рвота, диарея и головные боли. Вирус Тосканы, также переносимый москитами, обладает способностью проникать в нервную ткань и, вдобавок к вышеперечисленным симптомам, вызывает асептический менингит и менингоэнцефалит. Флебовирусы, переносимые клещами, например, вирус Бханджа, вирус тяжелой лихорадки с синдромом тромбоцитопении, или вирус Хартленд, вызывают серьезные вспышки инфекций среди людей [16].
Эти вирусы получили свое имя от латинского названия москитов (Phlebotominae), которые являются их основными переносчиками. Вирионы флебовирусов имеют диаметр 100-125 нанометров. Внутри вириона находятся три вирусных рибонуклеопротеина (вРНП), содержащих геномные сегменты, однако для вируса лихорадки долины Рифт (RVFV) было показано [17], что вирионы также могут содержать ещё три дополнительных вРНП, образованных цепочками антигеномных РНК, комплементарных геномным вирусным РНК. Рецептор-распознающий аппарат вирусов представлен гетеродимерами гликопротеинов Gn и Gc, которые организованным способом распределены по мембране вириона.
Структура генома флебовирусов
Геном флебовирусов как и других представителей семейства Bunyaviridae, включает три молекулы РНК: PHК L, РНК M, РНК S, имеющие на 5′- и 3′- концах уникальные для каждого геномного сегмента комплементарные последовательности. РНК L (–)полярности кодирует белок репликазы L. (–)РНК M кодирует предшественник гликопротеинов G1 и G2. (±)РНК S кодирует белок нуклеокапсида N на (–)полярном участке (ближе к 3′) и неструктурный белок NSs на (+)полярном участке (ближе к 5′) (рис. 2).

Рисунок 2. Схема структуры генома флебовирусов. Отмечены участки РНК, обладающие (–)- и (+)полярностью. Пунктирной линией обозначен сайт протеолиза белкового продукта.
NSs выполняет ряд функций, среди которых подавление индукции интерферона, усиление репликации и транскрипции вирусной РНК и определение круга хозяев [18]. NSs через цепочку белковых факторов способен приводить к инактивации противовирусной протеинкиназы R организма-хозяина [19].
Механизмы транскрипции и репликации РНК флебовирусов
Остановка транскрипции (–)участка РНК S определяется межгенным сигналом терминации. Похожие сигналы терминации находятся в 5′-концевой области РНК M и РНК L. В результате синтезируются кэпированные, но неполиаденилированные (и, следовательно, не такие стабильные, как клеточные мРНК) субгеномные РНК [18]. Также было показано [17], что в инфицированных клетках наблюдается ранняя экспрессия белка NSs, к тому же при детальном анализе состава вирионов обнаружили, что в вирусную частицу может упаковываться как три геномных цепи, так и еще три антигеномных цепи. Считается, что антигеномная РНК S присутствует в вирионе для осуществления ранней транскрипции мРНК, кодирующей NSs, поскольку этот неструктурный белок способен регулировать клеточные процессы, и чем раньше он начнёт работать в зараженной клетке, тем интенсивнее будет протекать вирусная инфекция.
Жизненный цикл флебовирусов
Жизненный цикл состоит из следующих стадий (рис. 3):

Рисунок 3. Схема, демонстрирующая основные этапы цикла флебовируса
Тосповирусы и тенуивирусы
Название рода Tospovirus происходит от сокращения названия вируса пятнистого увядания томатов (tomato spotted wilt virus, ТоSWV), впервые выделенного в 1930 году из зараженных растений томата. Этот вирус имеет очень широкий спектр хозяев и важное хозяйственное значение, борьба с ним ведется, в основном, за счет контроля численности трипсов.
Структура генома тосповирусов и тенуивирусов
Представители родов Тospovirus и Tenuivirus (тенуивирусы близки к тосповирусам, но не имеют липидной оболочки) являются единственными известными РНКвирусами растений с двусмысленным геномом [23]. Геном тосповирусов представлен тремя РНК-сегментами: большим, средним и малым (L, M, S). РНК L кодирует репликазу L. РНК S, подобно таковой у флебовирусов, кодирует белок нуклеокапсида N в (–)области и неструктурный белок NSs в (+)области. Эти области не пересекаются, они разделены межгенным некодирующим участком, содержащим сигналы терминации транскрипции. М-сегмент генома имеет принципиально отличную от РНК М флебовирусов структуру, являясь амбисенсной РНК. РНК М тосповирусов имеет область (–)полярности, в которой находится последовательность, кодирующая мРНК GnGc — предшественника поверхностных гликопротеинов, а также участок (+)полярности в 5′-области, кодирующий белок межклеточного транспорта NSm. Эти последовательности также разделены межгенным участком (рис. 4). Механизмы транскрипции и репликации РНК этих вирусов сходны с таковыми у флебовирусов [18].

Рисунок 4. Схема структуры генома тосповирусов. Отмечены участки РНК, обладающие (–)- и (+)полярностью. Пунктирной линией обозначен сайт протеолиза белкового продукта.
Отдельного внимания заслуживает неструктурный белок, закодированный в S-сегменте генома тосповирусов — NSs. Основной его функцией является супрессия противовирусного сайленсинга РНК, системы малых интерферирующих РНК [5], [24], распознающих вирусные РНК, что приводит к деградации последних [25]. Логично предположить, что синтез такого белка должен происходить как можно раньше, поэтому, возможно, по аналогии с белком NSs флебовирусов, ранняя транскрипция такой последовательности происходит в результате наличия в вирионе, помимо геномной цепи РНК S, еще и соответствующей ей антигеномной.
Вирусы с двусмысленным РНК-геномом из семейства Arenaviridae
Помимо семейства Bunyaviridae, амбисенсные РНК имеют представители семейства Arenaviridae. Аренавирусы являются таксономической группой вирусов позвоночных с сегментированным двусмысленным РНК-геномом. Вирусы, инфицирующие млекопитающих, определены в род Mammarenavirus, а заражающие рептилий — в роды Reptarenavirus и Hartmanivirus [26].
Вирионы аренавирусов, как и рассмотренных выше буньявирусов, плеоморфны, а их диаметр может варьировать от 40 до 200 нанометров в зависимости от вида, однако и частицы одного вида могут заметно различаться по размерам [27]. Границы вириона представлены липопротеидной оболочкой — производной клеточной мембраны, модифицированной равномерно распределёнными гликопротеиновыми комплексами (гетеродимерный гликопротеин GP1/GP2). Гликопротеины синтезируются в виде предшественника, который разрезается примерно пополам клеточной протеиназой на рецептор-распознающую субъединицу GP1 и трансмембранную субъединицу GP2. Последняя ответственна за слияние мембран при проникновении в цитоплазму [28]. В вирионе гликопротеины ассоциированны с лежащими на внутренней стороне мембраны молекулами матриксного белка Z, выстилающего внутреннюю поверхность мембраны, и белка нуклеокапсида N. Белок N способен связываться с РНК, распознавать кэп и ингибировать интерфероновый ответ. Структурный белок Z в клетке выполняет ряд функций (в том числе ингибирование трансляции клеточных мРНК и подавление апоптоза), являясь фактором созревания вирусных частиц (отвечает за инициацию сборки вирионов и за их отпочковывание).
Во время сборки вирусных частиц при формировании внешней оболочки иногда происходит захват субъединиц клеточных рибосом, по всей видимости, не играющих роли в вирусной инфекции (рис. 5).

Рисунок 5. Схематичное изображение структуры вириона аренавирусов
Значительная часть представителей семейства вызывает хронические и, как правило, бессимптомные инфекции у грызунов. При контакте человека с такими вирусами может развиваться острая и тяжелая инфекция, часто — геморрагическая лихорадка (например, в случае инфекции вирусом лихорадки Ласса, LasV). Вирус лимфоцитарного хориоменингита (LCMV), первый выделенный аренавирус, является нейроинвазивным. Попав в организм человека (например, через выделения грызунов), вирионы с током крови преодолевают гематоэнцефалический барьер центральной нервной системы и вызывают воспаления мозговых оболочек [29].
Структура генома аренавирусов

Рисунок 6. Схема структуры генома аренавирусов. Отмечены участки РНК, обладающие (–)- и (+)полярностью. Пунктирными линиями обозначены сайты протеолиза белковых продуктов.
РНК L на (–)участке несет последовательность, комплементарную гену репликазы L, и рамку матриксного и регуляторного белка Z на (+)участке в 5′-концевой части геномной РНК.
РНК S кодирует белок нуклеокапсида N в области (–)полярности и содержит рамку считывания GP1GP2 — предшественника поверхностных гликопротеинов GP1/GP2 (также в 5′-концевой части геномного сегмента).
Транскрипция и репликация генома аренавирусов
Переключение на репликацию связано с наличием белка N: когда его накапливается такое количество, что он начинает покрывать строящиеся цепи РНК, это, вероятно, влияет на конформацию репликазы и приводит к проскоку сигналов терминации транскрипции в виде межгенных шпилек. РНК S при репликации накапливается в больших количествах, так как нужно много копий белка нуклеокапсида, а также гликопротеинов (для экспрессии последних необходим предварительный синтез антигеномной РНК).
Заключение
Такая необычная организация двусмысленных геномных сегментов является интересным способом представления двух кодирующих последовательностей в одном геномном сегменте. На примере вирусов с двусмысленными РНК-геномами заметно, насколько изобретательной может быть эволюция вирусных РНК. Поскольку вирусы с двусмысленными РНК-геномами до сих пор удерживают определенную нишу, можно утверждать, что такой способ кодирования обладает некоторыми преимуществами по сравнению с более привычным для родственных вирусов способом, использующим только (–)РНК-сегменты.
Как возникли амбисенсные РНК и почему поддержались отбором, до сих пор остается одной из загадок современной вирусологии.
Благодарности от автора
Я благодарю доктора биологических наук, профессора кафедры вирусологии биологического факультета МГУ им. М.В. Ломоносова Аграновского Алексея Анатольевича за интересные и содержательные лекции по молекулярным процессам РНК-вирусов и вдохновение на написание данной статьи. Также выражаю благодарность художнику Андрею Занкевичу, чьи наглядные и яркие иллюстрации украшают данную статью.

Новость
Электронная микрофотография вирионов crAss-подобных фагов
Автор
Редактор
Тщательное изучение вирусных белков, даже самых многочисленных, подчас преподносит исследователям множество сюрпризов. Российские и американские ученые детально изучили РНК-полимеразу бактериофага φ14:2, входящего в семейство сrAss-подобных фагов — самой многочисленной группы вирусов в желудочно-кишечном тракте человека. Этот фермент упаковывается в вирионы вместе с вирусным геномом и участвует в транскрипции генов фага в начале инфекции. Исследователям удалось получить кристаллическую структуру РНК-полимеразы фага φ14:2, которая, как оказалось, очень похожа на эукариотические РНК-полимеразы, задействованные в РНК-интерференции. Авторы работы высказывают предположение, что эукариотические РНК-полимеразы, участвующие в РНК-интерференции, позаимствованы у фага, который, вероятно, инфицировал бактерию — предка митохондрий.
Чтобы совершить удивительное открытие в мире вирусов и микроорганизмов, вовсе не обязательно спускаться в океанские глубины, покорять горные вершины или забираться в другие труднодоступные места: зачастую необычные вещи спрятаны буквально у нас под носом. Так обстоят дела и с crAss-подобными бактериофагами , которые обитают в кишечнике каждого человека, но до сих пор оставались совершенно не изученными. Лишь в 2014 году, ученые, анализируя метагеном кишечного микробиома человека, обнаружили в нем последовательности ДНК, которые удалось собрать в геном длиной 100 тысяч пар оснований. Оказалось, что этот геном принадлежал ранее неизвестному бактериофагу, который получил название crAssphage (по названию метода, используемого для сборки генома — cross assembly) [1]. Выяснилось также, что crAssphage — это самый распространенный фаг в популяции человека. На его геном приходится до 90% всех ридов (прочтений), получаемых при секвенировании вирусной фракции микробиома кишечника человека [1]. Позже группа Евгения Кунина открыла, что в природе существует множество фагов, родственных crAss-фагу, но никто из них не был изучен в лабораторных условиях [2]. И это неудивительно, ведь большинство crAss-подобных фагов не культивировали в лаборатории, и мы знаем о них лишь из исследований метагеномов. Изучение crAss-подобных фагов представляет огромный интерес, поскольку они — самые многочисленные вирусы в кишечнике человека и, безусловно, оказывают влияние на наш микробиом . Кроме того, большинство генов crAss-подобных фагов мало на что похожи и кодируют новые, пока неизвестные белки.
Российские и американские ученые, среди которых специалисты Сколковского института науки и технологий, нашли crAss-подобный вирус — φ14:2, удобно культивируемый в лаборатории, и, используя его как модель, разобрались, каким образом crAss-подобные фаги транскрибируют свои гены [8]. Фаг φ14:2 поражает бактерию Cellulophaga baltica. Один из его генов кодирует большой белок gp66 (2180 аминокислотных остатков, а. о.), аминокислотная последовательность которого содержит маленький участок (всего 50 а. о.), похожий на фрагмент одной из субъединиц клеточных РНК-полимераз. Этот участок включает мотив, содержащий три абсолютно консервативных остатка аспартата, которые в многосубъединичных РНК-полимеразах координируют ионы магния и образуют каталитический центр фермента. Чтобы изучить функции gp66 и его ортологов в других crAss-подобных фагах, ученые экспрессировали ген белка gp66 в клетках кишечной палочки E. coli, очистили белок и показали, что он действительно может синтезировать РНК по матрице ДНК в присутствии ионов магния. И это несмотря на то, что gp66 похож на обычные РНК-полимеразы лишь своим крошечным фрагментом (менее 3% от всей величины белка). Вот уж поистине иголку в стоге сена нашли и предсказали биоинформатики!
Самый достоверный и точный способ узнать, как именно функционирует белок, — разрешить его пространственную структуру. Но для этого необходимо получить кристаллы белка, что часто становится весьма нетривиальной задачей, особенно для таких больших белков, как gp66. Однако исследователи смогли кристаллизовать gp66 и получить его структуру с разрешением в 3,5 ангстрем с помощью рентгеноструктурного анализа.
По структуре gp66 оказался наиболее близок к РНК-полимеразе QDE-1 грибка Neurospora crassa, которая участвует в синтезе малых интерферирующих РНК (рис. 1) [9]. Стоит, однако, отметить, что высокая степень сходства между gp66 и эукариотическими РНК-полимеразами, синтезирующими малые интерферирующие РНК, наблюдается лишь в областях белков, критически важных для выполнения основных функций. РНК-полимераза gp66 значительно больше, чем QDE-1, и имеет много дополнительных доменов с пока неизвестными функциями.

Рисунок 1. Структура gp66 фага φ14:2 (а) и РНК-полимеразы QDE-1 N. crassa [10] (б). Серым цветом окрашены различающиеся структурные элементы, одинаковыми цветами — аналогичные элементы структуры.
РНК-полимераза N. crassa, наиболее близкая к gp66, и ее ортологи широко распространены у современных эукариот и, вероятно, имелись у последнего их общего предка [11]. Эти белки первоначально были описаны как РНК-зависимые РНК-полимеразы, хотя в дальнейшем оказалось, что in vitro они транскрибируют однонитевую ДНК намного эффективнее, чем РНК [12]. Авторы исследования предполагают, что РНК-полимеразы этой группы появились у эукариот благодаря вирусу, инфицирующему бактериального предка митохондрий. К слову, похожий сценарий заимствования РНК-полимераз у вирусов уже был описан. Так, транскрипционный аппарат митохондрий, по всей вероятности, произошел от РНК-полимеразы T7-подобных фагов, не родственной клеточным многосубъединичным РНК-полимеразам и полимеразам crAss-подобных фагов.
Что еще интересного можно сказать про РНК-полимеразы crAss-подобных фагов? Судя по всему, это необычайно гибкие белки, способные к денатурации и структурным перестройкам. Так, при инфицировании бактериальной клетки gp66, состоящий из 2180 а. о., ухитряется протиснуться через канал в хвосте фага диаметром всего около 4 нм. Скорее всего, для этого РНК-полимераза gp66 полностью разворачивается. Попав в цитоплазму клетки-хозяина, gp66 должен приобрести свою нативную структуру. Интересно, что структура, которую получили исследователи, имеет конформацию каталитического центра, ранее не описанную для других РНК-полимераз, и в такой форме не может поддерживать катализ. Вероятно, это способ регулировать активность РНК-полимераз crAss-фагов на поздних стадиях инфекции, и переход в активную форму происходит при упаковке РНК-полимеразы в капсид или во время ее выхода в цитоплазму клетки.
Профессор Центра наук о жизни Мария Соколова, руководившая исследованием, отмечает:
«Это первая работа, где изучили, как происходит развитие инфекции crAss-подобным фагом. До сих пор были опубликованы работы, где изучали распространенность crAss-подобных фагов, их связь с какими-либо заболеваниями и т.п. В одной работе crAss-подобного фага выделили из фекалий человека, но не продвинулись дальше масс-спектрометрического анализа его вирионов. Мы же разобрались в том, как транскрибируются гены фагов этой группы, используя ф14:2 как модельный организм. Кроме того, полученная нами структура — это первая структура РНК-полимеразы, которая имеет общее происхождение с многосубъединичными РНК-полимеразами клеточных организмов, но настолько сильно от них отличается, что совершенно неясно, как она функционирует! Существует какой-то необычный механизм регуляции активности этой РНК-полимеразы, над определением которого мы сейчас работаем. Это очень интересно с точки зрения эволюции РНК-полимераз и процесса транскрипции в целом. Ну и наконец, РНК-полимераза вируса ф14:2 — это первый белок crAss-подобных фагов, для которого была экспериментально установлена функция. Бионформатические подходы, конечно же, очень важны и без них бы мы ничего не знали, но без эксперимента они дают не более чем предсказание.
ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
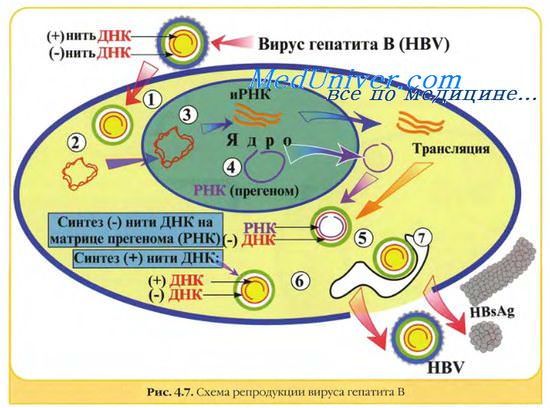
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Строение вирусов. Классификация вирусов
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
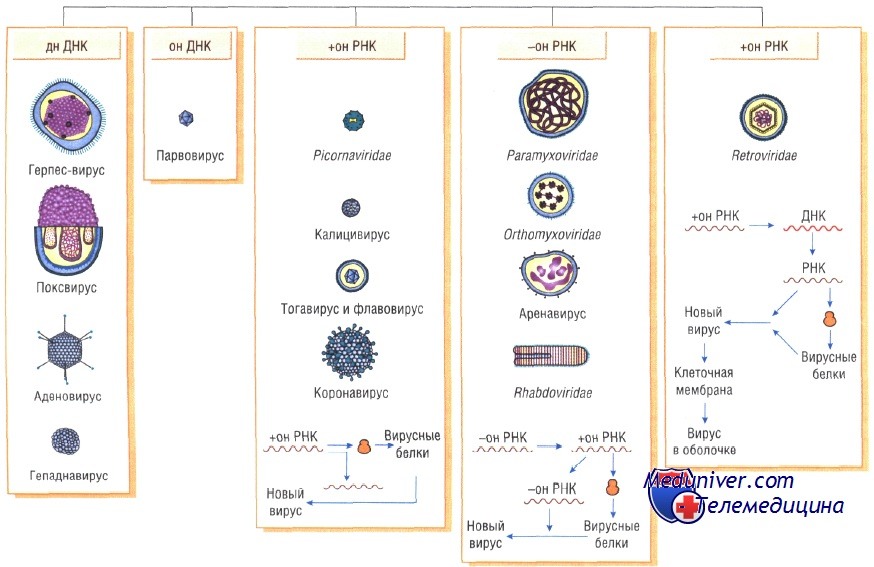
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
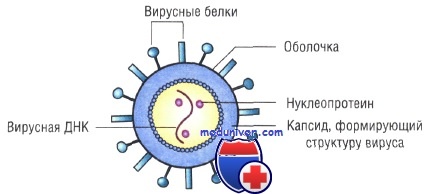
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: