
Вирусы могут репродуцироваться только
Обновлено: 25.04.2024
Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
Матричные белки необходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Типы инфицирования клеток вирусами. Репродуктивный цикл вирусов. Основные этапы репродукции вирусов. Адсорбция вириона к клетке.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Репродуктивный цикл вирусов
Изображённые на рис. 2-3 этапы репродукции (от адсорбции вирионов до высвобождения дочерней популяции) происходят при продуктивном взаимодействии вируса с клеткой.
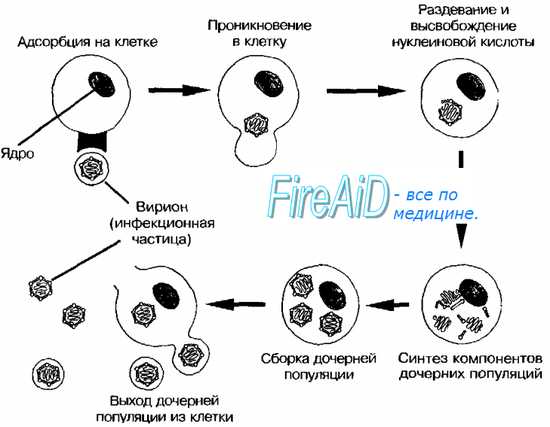
Рис. 2-3. Основные этапы репродукции вирусов.
Адсорбция вириона к клетке
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Репликация вирусов. Размножение вирусов.
Биотехнологии в вирусологии.
Методология выращивания вирусов.
Культивирование вирусов. Антигены вирусов и иммуннитет.
Иммунная система при вирусной инфекции.
Патогенез и механизмы противовирусной защиты организма.
Развитие и образование противовирусного иммунитета.

Вакцинопрофилактика вирусных инфекций.
Физические методы инактивации вирусов для вакцин.
Живые вакцины. Гетерологичные вакцины. Субъединичные вакцины.

Современные субъединированные и рекомбинантные вакцины.

Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Репродукция вируса – это процесс размножения вирусных частиц в чувствительных к ним клетках. Репродуцируются только вирулентные вирусы, обладающие высокой степенью патогенности [3] .
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций [3] .
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина [2] .
- Все вирусы, содержащие молекулу ДНК, кроме вирусаоспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирусоспы размножается только в цитоплазме клетки [2] .
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами [2] .
- Белки вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению) [2] .
- Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование (присоединении к полипептиду углеводных остатков) [2] .

Этапы репродукции (жизненного цикла) вируса

1. Адсорбция вируса на мембране клетки.; 2. Проникновение вируса в клетку.; 3. Депротеинизация.; 4. Синтез компонентов вирусов.; 5. Формирование дочерних вирионов.; 6. Выход вирионов [2] .
Этапы репродукции вируса
Репродукцию или жизненный цикл вируса делят на шесть последовательных этапов:
- Адсорбция на мембране клетки [2] .
- Проникновение в клетку [2] .
- Депротеинизация [2] .
- Синтез компонентов вирусов[2] .
- Формирование дочерних вирионов[2] .
- Выход вирионов[2] .

Процесс адсорбции вириона на поверхности клетки

1. Вирион.; 2. Клеточный рецептор.; 3. Прикрепительный белок; 4. Клеточная мембрана [2] .
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона [2] .
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур [2] .
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно [2] .
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 [2] .

Проникновение вируса в клетку (Путь I)

Слияние вирусной оболочки с клеточной мембраной
1. Вирион.; 2. Инфицируемая клетка.; 3. Ядро [2] .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона [2] . Существует два пути:
- Путь I – слиянии вирусной оболочки с клеточной мембраной [2] .
- Путь II – рецептор-опосредованный эндоцитоз [2] .
Путем слияния суперкапсида с клеточной мембраной (путь I) в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки [2] .
Путем рецептор-опосредованного эндоцитоза (путь II) в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении, вирусный геном в цитоплазме клетки освобождается из эндосомы [2] .

Проникновение вируса в клетку (Путь II)

1. Вирион.; 2. Клетка.; 3. Эндосома.; 4. Ядро [2] .
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы) [2] .
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона [2] .
Синтез компонентов вируса
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [1] .
Место синтеза компонентов дочерних вирионов зависит от типа генома:
- реализация генетической информации у ДНК-содержащих вирусов идет по пути: ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом идет по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок;
- РНК-содержащие ретровирусы идут по следующему пути передачи информации: РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок [2] .
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона [2] .
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках [2] .
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК [2] .
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается [2] .
РНК-содержащие ретровирусы отличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза [2] [4] .
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков, и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры [2] .
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более [2] .
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования[2] .
Путь лизиса клетки тесно связан ее деструкцией. Он характерен для безоболочечных вирусов, не имеющих суперкапсидной оболочки (суперкапсида) [2] .
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы, высвобождаются в течении 2–6 часов. В начале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида [2] .
Одни вирусы способны интегрироваться в геном клетки-мишени и таким образом оставаться во всех дочерних клетках, которые будут в будущем получены после ее деления. К таким вирусам относятся гаммаретровирусы и лентивирусы. Другие делать этого не умеют (например, адено- и аденоассоциированные вирусы). Но для производства белков и репликации (размножения) все они используют клетку и ее синтетический аппарат.
Как это работает?
Для того чтобы вирус мог проникнуть в клетку, белки его оболочки должны связаться с мембранными белками клетки-мишени. Важно отметить, что проникает вирус только в те клетки, которые могут в дальнейшем помочь его репликации. Вирус ВИЧ живет в клетках иммунной системы, вирус гепатита С — в клетках печени. Есть особые вирусы, которые поражают только растения или даже только бактерии.
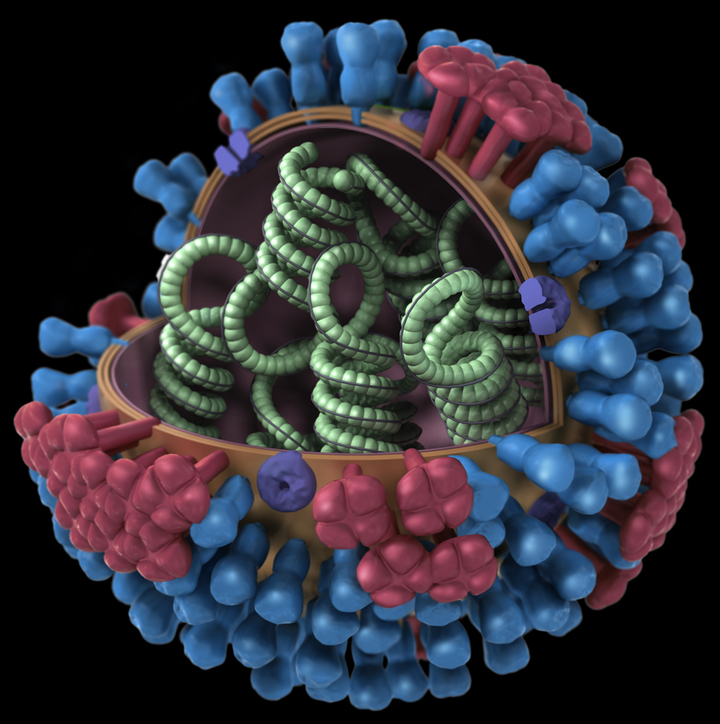
Строение вируса гриппа. Под оболочкой вириома - генетический материал вируса, необходимый для его воспроизводства в клетке.
Содержащие неактивный вирус и оставшиеся в живых клетки иногда сохраняют возможность нормального функционирования. В этом случае клетки могут быть заражены, но вирус проявит себя спустя длительный период времени. Так устроен герпес.
В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы. И тут стоит остановиться на классификации.
Типы вирусов. Коротко о главном
Современная типология вирусов содержит 7 классов и была предложена Дэвидом Балтимором еще в 1971 году. С тех пор, впрочем, она была уточнена и расширена, в том числе советскими учеными. И выглядит в настоящее время таким образом:
Вирусы, содержащие двухцепочечную ДНК
Описание
Для репликации вирусу необходимо попасть в ядро клетки-мишени и воспользоваться ее ДНК-полимеразой. Иногда вирус вызывает незапланированное деление самое клетки, то есть становится онкогенным. Эти вирусы хорошо изучены.
Пример: Вирус герпеса, адено- и папилломавирусы
Вирусы, содержащие одноцепочечную ДНК
Описание
Попадая в ядро клетки, вирусы образуют двухцепочечную ДНК, после чего реплицируются так же, как вирусы класса I.
Пример: Парво- и цирковирусы
Вирусы, в которых РНК способна к репликации (редупликации)
Описание
Вирусы этого класса могут размножаться в цитоплазме клетки, им не нужна молекула ДНК. Каждый ген, находящийся в РНК вируса, кодирует только один вирусный белок.
Пример: Бирна- и реовирусы
Вирусы, содержащие одноцепочечную (+) РНК
Описание
Из геномной (+) РНК на рибосомах хозяина создаются вирусные белки. В одном фрагменте РНК могут быть закодированы разные белки, что увеличивает сложность вируса без удлинения генов.
Пример: Пикорнавирусы (полиомиелит, гепатит А) и коронавирусы
Вирусы, содержащие одноцепочечную (–) РНК
Описание
(–) РНК этих вирусов предварительно должна быть транскрибирована в (+) РНК вирусными РНК-полимеразами, после чего может начаться синтез вирусных белков. Вирусы этого класса делятся еще на две группы, в зависимости от их генома и места его репликации (цитоплазма или ядро).
Пример: Филовирусы, аренавирусы (геморрагическая лихорадка Ласса), ортомиксовирусы (вирусы гриппа) и так далее.
Вирусы, содержащие одноцепочечную (+) РНК, реплицирующиеся через стадию ДНК
Описание
Такие вирусы используют фермент обратную транскриптазу для превращения (+) РНК в ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина.
Пример: Ретровирусы (в том числе ВИЧ)
Вирусы, содержащие двухцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК
Описание
Молекула ДНК замкнута в кольцо и является матрицей для синтеза мРНК и дополнительных молекул РНК, которые используются при репликации вирусного генома обратными транскриптазами.
Пример: Колимовирусы (вызывают инфекции растений) и гепаднавирусы (например, гепатит В)
Вакцинация и лечение
Как правило, организмы умеют бороться с паразитирующими на них вирусами. На примере млекопитающих и человека мы обычно говорим о главном инструменте — врожденном иммунитете.
Впрочем, наиболее эффективен этот вид защиты в отношении бактериальных инфекций и не может обеспечить продолжительную и надежную защиту, особенно от инфекций вирусных.
Именно поэтому огромное значение имеет приобретенный иммунитет, в результате которого клетки иммунной системы обучаются вырабатывать специфические к вирусу антитела, способные уничтожать как саму вирусную частицу, так и зараженные ею клетки.
Еще одна врожденная система борьбы с вирусными инфекциями — внутриклеточная. Как правило, клетка способна распознать чужеродную РНК в своей цитоплазме, куда ее сперва и доставляют многие вирусы, и имеет специальные комплексы для ее деградации. Но часть вирусов научились обходить и эту ловушку. К примеру, ротавирусы, которые даже внутри клетки сохраняют капсид с геномной РНК.
Миссия: уничтожить
Основная сложность в лечении вирусных заболеваний заключается в том, что они используют естественные функции клеток-мишеней для своего размножения, поэтому ученым зачастую оказывается не так-то просто придумать препарат, который будет токсичен для вируса и безопасен для самой клетки. Если такой безопасности достичь не удастся, лекарство будет иметь слишком много побочных эффектов, повреждающих сам организм, что окажется нецелесообразно для использования.

Сравнение жизненных циклов ВИЧ и вируса гриппа. Если первый использует обратную транскрипцию и живет в клетках иммунной стистемы, вирион второго, проникая в эпительные клетки дыхательных путей целиком - а именно там он и обитает - распадается уже внутри клетки, а репликация вирусной РНК происходит в ядре с помощью вирусных полимераз PA, PB1 и PB2 путем комплементарного копирования.
По принципу действия противовирусные препараты подразделяются на две группы: стимулирующие иммунную систему атаковать вирусы (например, за счет индукции синтеза белков-интерферонов) и атакующие вирусы напрямую. Препараты второй группы различаются по этапу жизненного цикла вируса, на котором они активны: это препараты, препятствующие проникновению вируса в клетку, препятствующие размножению вируса внутри клетки и препятствующие выходу копий вируса из клетки.
Еще один класс противовирусных препаратов блокирует ферменты, необходимые для создания и модификаций белков вируса. Такие лекарства называют протеазными ингибиторами.
Вместо заключения: а могут ли вирусы приносить пользу?
Безусловно, да. Несмотря на то, что вирусы ассоциируются у большинства людей с однозначным вредом, они могут приносить и пользу — если речь идет о так называемых вирусных векторах и терапевтических подходах на их основе.
Исследователи давно научились помещать в белковую оболочку вируса интересующие их нуклеиновые кислоты, чтобы доставлять нужный ген в клетки, а также убирать те гены, которые делают вирус опасным для организма.
Это позволило сделать возможной генную терапию, помогающую бороться с заболеваниями, вызванными известными генетическими мутациями. Создание вирусных векторов — достаточно непростая задача, к тому же ограниченная свойствами самих вирусных частиц: количеством помещающейся генетической информации, местом ее вставки, стабильностью. Кроме того, вирусный вектор, используемый в медицине, не должен вызывать иммунного ответа или критично влиять на жизнедеятельность клетки. Тем не менее эти сложности решаются, поэтому уже одобрен ряд вполне успешных и безопасных генных терапий. А в качестве основы для вирусных векторов чаще всего используются ретро-, ленти-, адено- и аденоассоциированные вирусы.
Читайте также: