
Заключение для реферата на тему онкогенные вирусы
Обновлено: 07.05.2024
Такая зависимость впервые была показана в 1908 г., когда датские ученые Эллерман и Банг нашли, что у цыплят возбудитель лейкоза может быть многократно пропущен через бактериальные фильтры и не теряет при этом своей агрессивности. В 1910 г. американец Пейтон Раус установил связь вирусов с саркомой кур. В 1934 г. его соотечественник Р. Шоуп выделил вирус из опухоли бородавок, или папиллом, кроликов. В 1936 г. Дж. Биттнером был открыт вирус рака молочной железы мышей, и 1951 г. Л. Гроссом — вирус лейкоза мышей, в 1964 г. В. Ярретом — вирус лейкоза домашних кошек, а затем и вирус лейкоза обезьян. В изучение последнего большой вклад внесли советские ученые под руководством академика АМН СССР Б. А. Лапина.
Содержание
Введение…………………………………………………………. 3-4
Особенности онкогенных вирусов…………………………..……4-6
Механизмы действия онковирусов……………………………. 6-9
Заключение………………………………………………………..9-11
Список использованной литературы…………………………. ….12
Вложенные файлы: 1 файл
онковирусы.doc
Министерство здравоохранения Республики Беларусь
Витебский государственный медицинский университет
Кафедра клинической микробиологии
Подготовила: студентка 3 к, 32 гр, л.ф.
Проверил: преподаватель кафедры
- Введение………………………………………………………… . 3-4
- Особенности онкогенных вирусов…………………………..……4-6
- Механизмы действия онковирусов……………………………. 6-9
- Заключение…………………………………………………… …..9-11
- Список использованной литературы…………………………. ….12
К числу факторов, которые наиболее достоверно способствуют трансформации протоонкоген а в онкоген, относятся некоторые вирусы. В 1907 г. французский исследователь А. Борель выдвинул гипотезу о связи рака с вирусами.
Такая зависимость впервые была показана в 1908 г., когда датские ученые Эллерман и Банг нашли, что у цыплят возбудитель лейкоза может быть многократно пропущен через бактериальные фильтры и не теряет при этом своей агрессивности. В 1910 г. американец Пейтон Раус установил связь вирусов с саркомой кур. В 1934 г. его соотечественник Р. Шоуп выделил вирус из опухоли бородавок, или папиллом, кроликов. В 1936 г. Дж. Биттнером был открыт вирус рака молочной железы мышей, и 1951 г. Л. Гроссом — вирус лейкоза мышей, в 1964 г. В. Ярретом — вирус лейкоза домашних кошек, а затем и вирус лейкоза обезьян. В изучение последнего большой вклад внесли советские ученые под руководством академика АМН СССР Б. А. Лапина.
Особенности онкогенных вирусов
РНК-содержащие: семейство Retroviridae.
ДНК-содержащие: семейства Papillomaviridae, Polyomaviridae, Adenoviridae 12, 18, 31, Hepadnaviridae, Herpesviridae, Poxviridae
Семейство Retroviridae включает 7 родов.
Онковирусы являются сложноорганизованными вирусами. Вирионы построены из сердцевины, окруженной липопротеиновой оболочкой с шипами. Размеры и формы шипов, а также локализация сердцевины служат основой для подразделения вирусов на 4 морфологических типа (А, В, С, D), а также вирус бычьего лейкоза.
Капсид онковирусов построен по кубическому типу симметрии. В него заключены нуклеопротеин и фермент ревертаза. Ревертаза обладает способностью транскрибировать ДНК. Геном – 2 идентичные цепи РНК.
В настоящее время известно более 30 видов опухолей млекопитающих, птиц и амфибий, которые вызываются вирусами, и более 150 онкогенных вирусов, способных их вызвать, Онкогенные вирусы обладают основным общим свойством — способностью трансформировать нормальные клетки в опухолевые. При проникновении вируса в клетку он не разрушает ее, а изменяет (трансформирует) в сторону беспрепятственного размножения, делая клетку злокачественной для организма. Онкогенными свойствами обладают различные представители как ДНК-, так и РНК-содержащих вирусов.
Установлена и достаточно изучена вирусная этиология доброкачественных опухолей — папиллом — у человека, возбудители которых относятся к семейству паповавириде. Оно состоит из трех родов: папилломавирус(па), полиомавирус(по) и вирусы вакуолизирующего агента (ва). Вирусы мелкие, размеры 43—55 нм (в среднем 30—35 нм), имеют молекулярную массу 3—5х16*6 дальтон, двунитчатую кольцевидную ДНК, составляющую 7—15% от массы вириона. Капсид голый, кубической симметрии, 72 капсомера. Созревание вируса происходит в ядре клетки.
Паповавирусы вызывают доброкачественные или злокачественные новообразования у животных. Папилломавирусы поражают эпидермис, приводят к возникновению доброкачественных папиллом кожи и слизистых оболочек (бородавки и кандиломы) у природных хозяев (кролики, золотистые хомяки, собаки, кошки, крупный рогатый скот, козы, обезьяны) и у человека.
Культивирование вирусов: не культивируются на куриных эмбрионах, культивируются в организме чувствительных животных, в культурах клеток.
Репродукция вирусов: проникают в клетку путем эндоцитоза. 3 этапа: синтез ДНК, на матрице РНК; ферментативное расщепление матричной РНК; синтез комплементарной нити ДНК на матрице первой нити ДНК.
К семейству Retroviridae относит ся примерно 150 видов вирусов, вызывающих развитие опухолей у животных, и только 4 вида вызывают опухоли у человека: HTLV-1, HTLV-2, ВИЧ-1,ВИЧ-2.
Вирусы Т-клеточного лейкоза человека
К семейству Retroviridae роду De ltaretrovirus относятся вирусы, поражающие CD4 Т-лимфоциты, для которых доказана этиологическая роль в развитии опухолевого процесса у людей: HTLV-1 и HTLV-2
Вирус HTLV-1 является возбудителем Т-клеточного лимфолейкоза взрослых. Он является экзогенным онковирусом, который, в отличие от других онковирусов, имеет два дополнительных структурных гена: tax и rех.
Продукт tax-гена действует на терминальные повторы LTR, стимулируя синтез вирусной иРНК, а также образование ИЛ-2 рецепторов на поверхности зараженной клетки. Продукт rex-гена определяет очередность трансляции вирусных иРНК.
HTLV-2 был изолирован от больного волосисто- клеточным лейкозом.
Оба вируса передаются половым, трансфузионным и трансплацентарным путями.
Семейство Papillomaviridae – вирус папилломы человека, собак. Вызывают инфекцию в клетках плоского эпителия. Доброкачественные папилломы в области половых органов, на коже, на слизистых дыхательных путей.
Семейство Polyomaviridae – вакуолизирующий вирус обезьян SV-40.Вирус полиомы человека.
Семейство Adenoviridae – аденовирусы, особенно серотипы 12,18,31 – индуцируют саркомы и трансформируют культуры клеток.
Семейство Poxviridae – вирусы фибромы-миксомы кролика, вирус Ябы, вызывающий развитие опухолей, вирус контагиозного моллюска.
Семейство Herpesviridae – лимфомы, карциномы. Онкогенез у человека связан с вирусом простого герпеса 2 типа (ВПГ-2) и вирусом Эпштейна-Барр (ВЭБ).
Новообразования, вызываемые онкогенными вирусами
У человека вирусы вызывают следующие новообразования.
1. Папилломы кожи и слизистых оболочек — разрастание покровного эпителия в виде сосочков над поверхностью кожи, слизистых оболочек рта, дыхательных и мочеполовых путей. Иногда они приобретают злокачественное течение. Вирусы человека отличаются от вирусов животных.
2. Инфекционные бородавки: обыкновенные плоские и остроконечные кондиломы, папилломы слизистых оболочек рта. В пораженных клетках обнаруживаются внутриядерные включения. Болеют чаще дети и юноши. Заражение происходит при прямом контакте с больными или через предметы общего пользования.
3. Контагиозный моллюск — мелкие плотные узелки у взрослых в области лобка, на половых органах, у детей на лице, веках, шее. При надавливании на них выделяется белая кашицеобразная масса с овоидными тельцами— моллюсковые тельца. Заражение происходит при контакте с больными, чаще половым путем.
Полиовирусы вызывают опухоли у природных хозяев: мышей, крыс, кроликов, хомяков, морских свинок и др., а также у обезьян макак резусов. Вирусы полиомы более мелкие и могут индуцировать злокачественные новообразования у новорожденных грызунов.
Среди РНК-содержащих вирусов имеются многочисленные опухолевые, онкогенные онкорнавирусы, вызывающие разнообразные опухоли. Все они объединены в семейство ретровириде, ранее род лейковирусов. Вирусы содержат однонитчатую РНК, молекулярная масса их 10—12ХІ06 дальтон, что составляет 1—2% от массы вириона. Тип симметрии возможно спиральный, число капсомеров неизвестно. Вирион покрыт оболочкой, чувствителен к эфиру. Диаметр вириона 100 нм, размножается в цитоплазме клеток. Вирусы не оказывают цитопатического действия на клетки. Основа группы онкорнавирусов (онко-опухоль, РНК-содержащий) —вирусы саркомно-лейкозного комплекса птиц и мышей, ответственны за естественно встречающиеся саркомы и лейкозы: вирус куриной саркомы Рауса, разнообразные штаммы вирусов лейкозов птиц и др. Предположительно считают возможным, что лейкозы человека вызывают вирус. Онкогенные вирусы в настоящее время разделяют на эндогенные и экзогенные. Эндогенные вирусы и их варианты открыты недавно, их находят в любых нормальных клетках и они не связаны с возникновением опухолей у природных хозяев, у которых они обнаруживаются. Однако, переходя на хозяина другого вида, они способны вызвать у него опухоль.
Механизмы действия онковирусов
Другой вариант: вирусный антиген заменяет один из белков тканевой совместимости I класса или заставляет их скучиваться и неправильно располагаться на мембране. В любом случае клетка должна быть распознана ЕК-клетками иммунного надзора или Т-лимфоцитами. Но если вирусный белок обладает иммуносупрессорными свойствами, такое незначительное изменение рисунка поверхности трансформированных вирусом клеток лимфоциты могут не заметить.
С точки зрения молекулярной биологии рак не столь уж разноликое заболевание. Один и тот же ген может быть активирован в любой ткани. При этом характер опухоли не зависит от типа онкогена. Он определяется лишь типом ткани. Если вирус поражает геном лейкоцитов — развивается лейкоз, при активации онкогена в клетках мышц или соединительной ткани возникает саркома, трансформация эпителиальных клеток влечет за собой рак.
До недавнего времени оставалось много сомнений относительно того, являются ли вирусы причиной опухолей только у животных или существуют опухолеродные вирусы человека. Постепенно накапливалось немало косвенных свидетельств участия вирусов в онкологических заболеваниях человека. Так, рак печени часто развивается у людей, переболевших инфекционным гепатитом В (болезнью Боткина). Эпидемиологические и иммунологические исследования показали, что почти 90% больных первичным раком печени имеют антитела к вирусу В или содержат его частицы в клетках опухоли.
В 1958 г. английский хирург Д. Беркит описал особо распространенную в Экваториальной Африке злокачественную опухоль лица, поражающую преимущественно детей. Основную часть опухолевой массы составляли лимфоидные клетки, поэтому болезнь получила название лимфомы Беркита. В дальнейшем было показано, что наиболее вероятным возбудителем ее является один из вирусов группы герпеса, названный вирусом Эпштейн—Барра в честь английских вирусологов, открывших его. Вирус этот распространен относительно широко, а лимфома Беркита является редким заболеванием (в Африке регистрируется 5—10 тыс. случаев в год). Далее оказалось, что с тем же вирусом может быть связан рак носоглотки, который особо часто встречается в Южной Азии, на юге Китая, в Северной Африке и у эскимосов. Кроме того, вирус Эпштейн—Барра считают основным виновником инфекционного мононуклеоза, при котором отмечается разрастание лимфоидной ткани, преходящее увеличение лимфатических узлов и увеличение числа незрелых лимфоцитов в крови.
В последние годы М. Эпштейн с сотрудниками получили два варианта вакцины против вируса Эпштейн— Барра. Один вариант представляет искусственные пузырьки из липопротеина-липосомы, начиненные гликопротеином наружных мембран вируса, другой — сами мембраны инфицированных вирусом клеток человека. Оба типа вакцин связывали антитела к вирусу и предупреждали у обезьян развитие вирус-индуцированного лейкоза.
У большинства, если не у всех женщин, больных раком шейки матки и наружных половых органов, находят вирусы папилломатоза или антитела к ДНК некоторых вирусов этого семейства. Внедрение вирусной ДНК в геном клеток может быть первым шагом на пути к малигнизации, поэтому папилломатозный вирус, особенно его типы 16 и 18, считают потенциальными онкогенами. Эти вирусы содержатся в клетках длительно перевиваемой культуры HeLa. Если к ним добавить нормальные клетки, то и те в скором времени трансформируются в опухолевые. Интересно, что при трансплантации таких неотрансформированных клеток мышам-нюдам опухоли у реципиентов не развиваются; под действием неизвестных антагонистов вирусный геном устраняется.
Такая зависимость впервые была показана в 1908 г., когда датские ученые Эллерман и Банг нашли, что у цыплят возбудитель лейкоза может быть многократно пропущен через бактериальные фильтры и не теряет при этом своей агрессивности. В 1910 г. американец Пейтон Раус установил связь вирусов с саркомой кур. В 1934 г. его соотечественник Р. Шоуп выделил вирус из опухоли бородавок, или папиллом, кроликов. В 1936 г. Дж. Биттнером был открыт вирус рака молочной железы мышей, и 1951 г. Л. Гроссом — вирус лейкоза мышей, в 1964 г. В. Ярретом — вирус лейкоза домашних кошек, а затем и вирус лейкоза обезьян. В изучение последнего большой вклад внесли советские ученые под руководством академика АМН СССР Б. А. Лапина.
Содержание
Введение…………………………………………………………. 3-4
Особенности онкогенных вирусов…………………………..……4-6
Механизмы действия онковирусов……………………………. 6-9
Заключение………………………………………………………..9-11
Список использованной литературы…………………………. ….12
Вложенные файлы: 1 файл
онковирусы.doc
Министерство здравоохранения Республики Беларусь
Витебский государственный медицинский университет
Кафедра клинической микробиологии
Подготовила: студентка 3 к, 32 гр, л.ф.
Проверил: преподаватель кафедры
- Введение………………………………………………………… . 3-4
- Особенности онкогенных вирусов…………………………..……4-6
- Механизмы действия онковирусов……………………………. 6-9
- Заключение…………………………………………………… …..9-11
- Список использованной литературы…………………………. ….12
К числу факторов, которые наиболее достоверно способствуют трансформации протоонкоген а в онкоген, относятся некоторые вирусы. В 1907 г. французский исследователь А. Борель выдвинул гипотезу о связи рака с вирусами.
Такая зависимость впервые была показана в 1908 г., когда датские ученые Эллерман и Банг нашли, что у цыплят возбудитель лейкоза может быть многократно пропущен через бактериальные фильтры и не теряет при этом своей агрессивности. В 1910 г. американец Пейтон Раус установил связь вирусов с саркомой кур. В 1934 г. его соотечественник Р. Шоуп выделил вирус из опухоли бородавок, или папиллом, кроликов. В 1936 г. Дж. Биттнером был открыт вирус рака молочной железы мышей, и 1951 г. Л. Гроссом — вирус лейкоза мышей, в 1964 г. В. Ярретом — вирус лейкоза домашних кошек, а затем и вирус лейкоза обезьян. В изучение последнего большой вклад внесли советские ученые под руководством академика АМН СССР Б. А. Лапина.
Особенности онкогенных вирусов
РНК-содержащие: семейство Retroviridae.
ДНК-содержащие: семейства Papillomaviridae, Polyomaviridae, Adenoviridae 12, 18, 31, Hepadnaviridae, Herpesviridae, Poxviridae
Семейство Retroviridae включает 7 родов.
Онковирусы являются сложноорганизованными вирусами. Вирионы построены из сердцевины, окруженной липопротеиновой оболочкой с шипами. Размеры и формы шипов, а также локализация сердцевины служат основой для подразделения вирусов на 4 морфологических типа (А, В, С, D), а также вирус бычьего лейкоза.
Капсид онковирусов построен по кубическому типу симметрии. В него заключены нуклеопротеин и фермент ревертаза. Ревертаза обладает способностью транскрибировать ДНК. Геном – 2 идентичные цепи РНК.
В настоящее время известно более 30 видов опухолей млекопитающих, птиц и амфибий, которые вызываются вирусами, и более 150 онкогенных вирусов, способных их вызвать, Онкогенные вирусы обладают основным общим свойством — способностью трансформировать нормальные клетки в опухолевые. При проникновении вируса в клетку он не разрушает ее, а изменяет (трансформирует) в сторону беспрепятственного размножения, делая клетку злокачественной для организма. Онкогенными свойствами обладают различные представители как ДНК-, так и РНК-содержащих вирусов.
Установлена и достаточно изучена вирусная этиология доброкачественных опухолей — папиллом — у человека, возбудители которых относятся к семейству паповавириде. Оно состоит из трех родов: папилломавирус(па), полиомавирус(по) и вирусы вакуолизирующего агента (ва). Вирусы мелкие, размеры 43—55 нм (в среднем 30—35 нм), имеют молекулярную массу 3—5х16*6 дальтон, двунитчатую кольцевидную ДНК, составляющую 7—15% от массы вириона. Капсид голый, кубической симметрии, 72 капсомера. Созревание вируса происходит в ядре клетки.
Паповавирусы вызывают доброкачественные или злокачественные новообразования у животных. Папилломавирусы поражают эпидермис, приводят к возникновению доброкачественных папиллом кожи и слизистых оболочек (бородавки и кандиломы) у природных хозяев (кролики, золотистые хомяки, собаки, кошки, крупный рогатый скот, козы, обезьяны) и у человека.
Культивирование вирусов: не культивируются на куриных эмбрионах, культивируются в организме чувствительных животных, в культурах клеток.
Репродукция вирусов: проникают в клетку путем эндоцитоза. 3 этапа: синтез ДНК, на матрице РНК; ферментативное расщепление матричной РНК; синтез комплементарной нити ДНК на матрице первой нити ДНК.
К семейству Retroviridae относит ся примерно 150 видов вирусов, вызывающих развитие опухолей у животных, и только 4 вида вызывают опухоли у человека: HTLV-1, HTLV-2, ВИЧ-1,ВИЧ-2.
Вирусы Т-клеточного лейкоза человека
К семейству Retroviridae роду De ltaretrovirus относятся вирусы, поражающие CD4 Т-лимфоциты, для которых доказана этиологическая роль в развитии опухолевого процесса у людей: HTLV-1 и HTLV-2
Вирус HTLV-1 является возбудителем Т-клеточного лимфолейкоза взрослых. Он является экзогенным онковирусом, который, в отличие от других онковирусов, имеет два дополнительных структурных гена: tax и rех.
Продукт tax-гена действует на терминальные повторы LTR, стимулируя синтез вирусной иРНК, а также образование ИЛ-2 рецепторов на поверхности зараженной клетки. Продукт rex-гена определяет очередность трансляции вирусных иРНК.
HTLV-2 был изолирован от больного волосисто- клеточным лейкозом.
Оба вируса передаются половым, трансфузионным и трансплацентарным путями.
Семейство Papillomaviridae – вирус папилломы человека, собак. Вызывают инфекцию в клетках плоского эпителия. Доброкачественные папилломы в области половых органов, на коже, на слизистых дыхательных путей.
Семейство Polyomaviridae – вакуолизирующий вирус обезьян SV-40.Вирус полиомы человека.
Семейство Adenoviridae – аденовирусы, особенно серотипы 12,18,31 – индуцируют саркомы и трансформируют культуры клеток.
Семейство Poxviridae – вирусы фибромы-миксомы кролика, вирус Ябы, вызывающий развитие опухолей, вирус контагиозного моллюска.
Семейство Herpesviridae – лимфомы, карциномы. Онкогенез у человека связан с вирусом простого герпеса 2 типа (ВПГ-2) и вирусом Эпштейна-Барр (ВЭБ).
Новообразования, вызываемые онкогенными вирусами
У человека вирусы вызывают следующие новообразования.
1. Папилломы кожи и слизистых оболочек — разрастание покровного эпителия в виде сосочков над поверхностью кожи, слизистых оболочек рта, дыхательных и мочеполовых путей. Иногда они приобретают злокачественное течение. Вирусы человека отличаются от вирусов животных.
2. Инфекционные бородавки: обыкновенные плоские и остроконечные кондиломы, папилломы слизистых оболочек рта. В пораженных клетках обнаруживаются внутриядерные включения. Болеют чаще дети и юноши. Заражение происходит при прямом контакте с больными или через предметы общего пользования.
3. Контагиозный моллюск — мелкие плотные узелки у взрослых в области лобка, на половых органах, у детей на лице, веках, шее. При надавливании на них выделяется белая кашицеобразная масса с овоидными тельцами— моллюсковые тельца. Заражение происходит при контакте с больными, чаще половым путем.
Полиовирусы вызывают опухоли у природных хозяев: мышей, крыс, кроликов, хомяков, морских свинок и др., а также у обезьян макак резусов. Вирусы полиомы более мелкие и могут индуцировать злокачественные новообразования у новорожденных грызунов.
Среди РНК-содержащих вирусов имеются многочисленные опухолевые, онкогенные онкорнавирусы, вызывающие разнообразные опухоли. Все они объединены в семейство ретровириде, ранее род лейковирусов. Вирусы содержат однонитчатую РНК, молекулярная масса их 10—12ХІ06 дальтон, что составляет 1—2% от массы вириона. Тип симметрии возможно спиральный, число капсомеров неизвестно. Вирион покрыт оболочкой, чувствителен к эфиру. Диаметр вириона 100 нм, размножается в цитоплазме клеток. Вирусы не оказывают цитопатического действия на клетки. Основа группы онкорнавирусов (онко-опухоль, РНК-содержащий) —вирусы саркомно-лейкозного комплекса птиц и мышей, ответственны за естественно встречающиеся саркомы и лейкозы: вирус куриной саркомы Рауса, разнообразные штаммы вирусов лейкозов птиц и др. Предположительно считают возможным, что лейкозы человека вызывают вирус. Онкогенные вирусы в настоящее время разделяют на эндогенные и экзогенные. Эндогенные вирусы и их варианты открыты недавно, их находят в любых нормальных клетках и они не связаны с возникновением опухолей у природных хозяев, у которых они обнаруживаются. Однако, переходя на хозяина другого вида, они способны вызвать у него опухоль.
Механизмы действия онковирусов
Другой вариант: вирусный антиген заменяет один из белков тканевой совместимости I класса или заставляет их скучиваться и неправильно располагаться на мембране. В любом случае клетка должна быть распознана ЕК-клетками иммунного надзора или Т-лимфоцитами. Но если вирусный белок обладает иммуносупрессорными свойствами, такое незначительное изменение рисунка поверхности трансформированных вирусом клеток лимфоциты могут не заметить.
С точки зрения молекулярной биологии рак не столь уж разноликое заболевание. Один и тот же ген может быть активирован в любой ткани. При этом характер опухоли не зависит от типа онкогена. Он определяется лишь типом ткани. Если вирус поражает геном лейкоцитов — развивается лейкоз, при активации онкогена в клетках мышц или соединительной ткани возникает саркома, трансформация эпителиальных клеток влечет за собой рак.
До недавнего времени оставалось много сомнений относительно того, являются ли вирусы причиной опухолей только у животных или существуют опухолеродные вирусы человека. Постепенно накапливалось немало косвенных свидетельств участия вирусов в онкологических заболеваниях человека. Так, рак печени часто развивается у людей, переболевших инфекционным гепатитом В (болезнью Боткина). Эпидемиологические и иммунологические исследования показали, что почти 90% больных первичным раком печени имеют антитела к вирусу В или содержат его частицы в клетках опухоли.
В 1958 г. английский хирург Д. Беркит описал особо распространенную в Экваториальной Африке злокачественную опухоль лица, поражающую преимущественно детей. Основную часть опухолевой массы составляли лимфоидные клетки, поэтому болезнь получила название лимфомы Беркита. В дальнейшем было показано, что наиболее вероятным возбудителем ее является один из вирусов группы герпеса, названный вирусом Эпштейн—Барра в честь английских вирусологов, открывших его. Вирус этот распространен относительно широко, а лимфома Беркита является редким заболеванием (в Африке регистрируется 5—10 тыс. случаев в год). Далее оказалось, что с тем же вирусом может быть связан рак носоглотки, который особо часто встречается в Южной Азии, на юге Китая, в Северной Африке и у эскимосов. Кроме того, вирус Эпштейн—Барра считают основным виновником инфекционного мононуклеоза, при котором отмечается разрастание лимфоидной ткани, преходящее увеличение лимфатических узлов и увеличение числа незрелых лимфоцитов в крови.
В последние годы М. Эпштейн с сотрудниками получили два варианта вакцины против вируса Эпштейн— Барра. Один вариант представляет искусственные пузырьки из липопротеина-липосомы, начиненные гликопротеином наружных мембран вируса, другой — сами мембраны инфицированных вирусом клеток человека. Оба типа вакцин связывали антитела к вирусу и предупреждали у обезьян развитие вирус-индуцированного лейкоза.
У большинства, если не у всех женщин, больных раком шейки матки и наружных половых органов, находят вирусы папилломатоза или антитела к ДНК некоторых вирусов этого семейства. Внедрение вирусной ДНК в геном клеток может быть первым шагом на пути к малигнизации, поэтому папилломатозный вирус, особенно его типы 16 и 18, считают потенциальными онкогенами. Эти вирусы содержатся в клетках длительно перевиваемой культуры HeLa. Если к ним добавить нормальные клетки, то и те в скором времени трансформируются в опухолевые. Интересно, что при трансплантации таких неотрансформированных клеток мышам-нюдам опухоли у реципиентов не развиваются; под действием неизвестных антагонистов вирусный геном устраняется.

Инфекция, вызванная онкогенными вирусами, является причиной 15,4% заболеваний раком по всей земле. Первый онкогенный вирус, который был идентифицирован — это куриный вирус саркомы Рауса в 1911 году. Десятилетиями позже были обнаружены серии других онкогенных вирусов, в том числе вирус папилломы Шопа (вирус папилломы кроличьего хвоста), вирус опухоли молочной железы мыши, аденовирус, вирус SV40 (обезьяний вирус sv40). Теория опосредованного вирусом онкогенеза в итоге была экспериментально продемонстрирована в 1976 году Гарольдом Вармусом и Майклом Бишопом путем превращения инфицированных клеток в опухолевые вирусом саркомы (ген v-src вызывает образование опухоли после захвата протоонкогена человеческой клетки в результате трансдукции).
Несмотря на то, что онкогенные вирусы нашли в исследованиях онкогенной активности животных клеток, они были не в состоянии преобразовать человеческие клетки. Теория вирусного онкогенеза в человеческой клетке оставалась спорной до 1965 года, когда вирус Эпштейн-Барр (EBV) был обнаружен в клетках лимфомы Беркитта. В ходе последующих исследований, в том числе выделении Т-лимфотропного вируса человека (HTLV) из клеток Т-клеточной лимфомы и выявлении роли папилломовирусов человека с высокой онкогенной активностью в развитии рака шейки матки, был определен путь для понятия человеческих онкогенных вирусов. Обнаружение связи между герпес-вирусом и саркомой Капоши (KSHV) и лимфомой и между полиомавирусом клеток Меркеля (MCV) и карциномой клеток Меркеля (MCC) подчеркнуло возможность открытия еще большего количества онкогенных вирусов с помощью современных технологий. Далее каждый вирус будет рассмотрен отдельно.
Вирус Эпштейн-Барр
Вирус Эпштейн-Барр (ВЭБ, EBV) — ДНК-содержащий вирус, который преимущественно поражает B-лимфоциты, также является митогеном для них; вызывает лимфопролиферативные болезни (лимфома Беркитта, носоглоточная карцинома, NK-клеточная лейкемия, лимфома Ходжкина и т.д.), а также инфекционный мононуклеоз. Вирионы вируса были обнаружены при электронной микроскопии биоптата лимфомы Беркитта в начале 1964 года Энтони Эпштейном и Ивонной Барр.
Строение. ВЭБ имеет вирусный капсидный антиген (VCA), ядерные антигены-EBNAs 1, 2, 3, 3a, 3b, 3c; латентные мембранные протеины (LMPs) 1, 2 и две маленькие Эпштейна-Барр-кодируемые РНК (EBER) молекулы — EBER1 и EBER2. EBNAs и LMPs являются ДНК-связывающими белками, считающимися необходимыми для развития инфекции и фиксации.
Промотор репликации плазмидного вектора показан оранжевым. Короткие толстые зеленые стрелки представляют экзоны, кодирующие латентные белки: (EBNAs 1, 2, 3A, 3B and 3C, and EBNA‑LP, LMPs 1, 2A and 2B, BHRF1,BARF1). Короткие голубые стрелки сверху представляют наиболее транскрибириуемые полиаденилированные ВЭБ-кодируемые РНК (EBER1, EBER2). Средняя длинная зеленая линия представляет транскрипцию ВЭБ при 3-ем типе латентности, в которой белки EBNA транскрибированы с промотора Ср или Wp. Различные EBNA кодированы индивидуальными мРНК, которые сгенерированы различным сплайсингом одного и того же длинного первичного транскрипта. Внутренняя красная линия представляет транскрипт EBNA1, который берет начало от промотора Qp при 1 и 2 типе латентности.
- необходим для трансформации В-лимфоцитов;
- функционирует как конститутивный активатор рецептора ФНО и имеет сходство с CD-40 рецептором;
- активирует сигнальные пути, которые делают вклад в онкогенный эффект.
В зависимости от набора экспрессируемых вирусных генов в пораженных клетках выделяют три типа латентности (I, II и III тип), характерные для различных видов лимфом: I тип подразумевает избирательную экспрессию EBNA-1 при ВЭБ-положительной лимфоме Беркитта; II тип заключается в экспрессии EBNA-1, LMP-1 и LMP-2 и является отличительным признаком ВЭБ-положительной ходжкинской лимфомы, периферических T/NK-клеточных лимфом; латентность III типа, характеризуемая экспрессией всего набора из девяти латентных белков ВЭБ, наблюдается при лимфопролиферативных поражениях, возникающих у пациентов с тяжелым иммунодефицитом (после трансплантации паренхиматозных органов или стволовых клеток, ВИЧ-инфекции). Тип латентности определяет восприимчивость инфицированных клеток к различным иммунотерапевтическим тактикам. Индуцированные ВЭБ злокачественные новообразования связаны с экспрессией латентных генов. У большинства иммунокомпетентных носителей ВЭБ вирус не индуцирует опухолевый процесс. Новообразования развиваются при комплексном взаимодействии ВЭБ, иммуногенетических факторов, факторов внешней среды (паразитарных инфекций — чаще, малярии, недоедании, потреблении пищи, содержащей канцерогены) и иммунодефицита (ВИЧ-инфекция, трансплантация).
Папилломавирусы человека
Описано около 200 папилломавирусов. Вирусы передаются при контактах через микротравмы кожи и слизистых оболочек, а также половым путем. Папилломавирусы человека вызывают доброкачественные кожные, генитальные, оральные и конъюнктивальные папилломы; индуцируют пролиферацию эпителия, обладают онкогенным потенциалом. С папилломавирусом, особенно ПВЧ-5 и ПВЧ-8, тесно связано развитие немеланомных разновидностей рака кожи. Различают низкоонкогенные папилломавирусы, вызывающие бородавки, которые не малигнизируются, и высокоонкогенные, способные вызвать рак шейки матки, — ПВЧ-16, 18, 31, 33, 35, 45, 51, 52, 58.
Строение. Капсид икосаэдрический, состоит из двух капсидных белков (L1-L2), формирующих 72 пентамера. Геном — двунитевая циркулярная ДНК; имеет в зависимости от вируса восемь ранних генов (E1-E8) и два поздних гена (L1-L2). Гены E6-E7 обладают онкогенной активностью.
Патогенез. В целом, для развития продуктивного поражения ВПЧ (высокого или низкого риска) требуется, чтобы вирус имел доступ к эпителию базального слоя как только эпителиальный барьер был подвержен риску. Папилломавирусы должны инфицировать делящуюся клетку, чтобы стать признанными как малое количество копий ядерной эписомы, и для развития стойкого поражения первичная инфицированная клетка, вероятно, должна быть похожа на долгоживующую эпителиальную стволовую клетку или похожую на нее. Кроме того, связанные с заживлением ран изменения в местном микроокружении, включая увеличение количества факторов роста, могут играть ключевую роль в создании резервуара инфекции в базальном слое. Способность ВПЧ инфицировать, и реплицироваться, и управлять прогрессированием рака зависит от функции вирусного белка, положении в эпителии, где эти белки экспрессируются. Как только инфицированные базальные клетки поделились и в конечном итоге вошли в парабазальный слой, они начинают проходить программу экспрессии генов, которая связана с дифференциацией. Амплификация вирусного генома обычно происходит в срединно-эпителиальных слоях, тогда как сбор и выход вируса происходит вблизи наружной поверхности эпителия.
Несмотря на то, что эффективная амплификация генома требует комбинированного действия множественных продуктов вирусного гена, включая E6, E7, E2 и кодированную вирусом геликазы E1, поддержание репликации в базальном слое эпителия может зависеть от факторов репликации клетки хозяина. Программа, зависящая от дифференциации, которая происходит в надбазальных слоях эпителиальной клетки, включает как временные изменения в активности промотора, так и изменения в шаблонах сплайсинга мРНК, которые облегчают производство различных продуктов вирусных генов на разных стадиях во время программы дифференциации. Функция вирусного белка далее регулируется посттрансляционной модификацией, включая фосфорилирование и протеолитическое деление клетки.

Герпесвирус, ассоциированный с саркомой Капоши
KSHV, или человеческий герпесвирус 8, был обнаружен в 1994 году как член семейства гамма-герпеса человека, присоединившись к ВЭБ. Инфекция иммунокомпрометированных индивидуумов с KSHV была связана с развитием саркомы Капоши, полученной из эндотелиальных клеток, и по меньшей мере двумя B-лимфоцит лимфопролиферативными заболеваниями: первичная выпотная лимфома и многоцентричная болезнь Кастлмана. Однако изучение патогенеза и онкогенеза KSHV затруднено отсутствием значимой модели животных и восприимчивой клеточной культуры.
Онкогенное действие. Две иммортализованные клеточные линии, KS Y-1 и SLK, когда-то использовались для исследований KS и KSHV, но в KS Y-1 были также обнаружены клетки линии рака мочевого пузыря T 24, что затруднило дальнейшую работу. Первичные эмбриональные метанефрические мезенхимальные клетки-предшественники почек крысы восприимчивы к инфекции и трансформации KSHV, но из них распространяются ограниченное количество инфекционных вирионов. Линии B-клеток, полученные из первичной выпотной лимфомы, обычно заражаются KSHV на латентной стадии и могут быть индуцированы для получения малого количества вирионов KSHV, но первичные B-лимфоциты из периферической крови или тонзиллярной ткани являются устойчивыми к инфекции KSHV, и для их инфицирования может потребоваться совместный рост с KSHV-позитивными клетками.
KSHV кодирует несколько важных белков, которые обладают некоторой онкогенной активностью для индуцирования пролиферации клеток, иммортализации, трансформации и передачи сигналов; производства цитокинов; иммунного контроля; антиапоптозной активности; и ангиогенеза. К ним относятся вирусные латентные белки, латентный ассоциированный ядерный антиген (LANA), vFLIP (FADD (Fas-ассоциированный белок с доменом смерти), подобный ферменту, ингибирующему синтез интерферона или ингибирующему белок каспазы 8 (FLICE)), vCyclin и вирусные литические белки G-белкового рецептора (vGPCR), интерферон-регуляторный фактор 1 (vIRF-1) и K1. Хотя истинную онкогенную природу каждого белка еще предстоит определить, накопление доказательств указывает на то, что каждый из них вносит свой вклад в онкогенез KSHV. Таким образом, полный спектр вызванной KSHV злокачественности может потребовать, чтобы несколько онкогенных продуктов работали вместе в присутствии клеток хозяев и экологических факторов. Например, как LANA, так и vIRF-1 нацелены на клеточный опухолевый супрессор p53. LANA также ингибирует pRB и PP2A.vCyclin, активатор CDK4 6,198, подавляет p27kip1, ингибитор CDK199 и вызывает реакцию на задержку старения G1, результаты от гиперактивации NF-κB. vFLIP и K1 активируют путь сигнала NF-κB для предотвращения апоптоза B-клеток. vGPCR и K1 влияют на сигнальные пути AKT и NF-κB и способствуют ангиопролиферативному и воспалительному поражению саркомой Капоши.
Т-лимфотропный вирус человека (HTLV)
T-лимфотропный вирус человека (HTLV) относится к семейству ретровирусов, является возбудителем T-клеточного лимфолейкоза взрослых. Вирус впервые был изолирован в 1980 году от больного Т-лимфомой.
Геном. Gag, Pol и Env являются вирусными структурными белками, другие — вирусными регуляторными/вспомогательными белками. За исключением гена hbz, который кодируется минус-цепью провирусного генома HTLV из 3'-LTR, все остальные гены кодируются плюсовой нитью по направлению 5'-LTR. Следует отметить, что 5'-LTR часто удаляется или метилируется по мере прогрессирования заболевания. Кроме того, на поздних стадиях лейкемиогенеза в гене tax часто возникают нонсенс- или миссенс-мутации. Хотя белок tax и ген HBZ индуцируют опухоли у трансгенных мышей, а p12 проявляет слабую онкогенную активность, ни один из вирусных белков/генов, кроме tax, не требуется для опосредованного HTLV-1 опухолеобразования. Продукт tax гена действует на терминальные повторы LTR, стимулируя синтез вирусной иРНК, а также образование рецепторов IL-2 на поверхности зараженной клетки.

Рисунок 3. Gag, Pol и Env вирусные регуляторные/вспомогательные белки HTLV

Рисунок 4. К леточные пути вирусного онкобелка tax
Патогенез. Проникновение HTLV-1 в клетку человека охарактеризовывает начало выживания и репликации вируса. HTLV-1 связывается с рецептором клеток, имеющих необходимый антиген, что инициирует процесс инвазии. Рецепторный комплекс состоит из транспортера глюкозы (GLUT1), гепаринсульфат протеогликана (HSPG) и рецептора VEGF-165 нейропилина-1 (NRP-1). HTLV-1 взаимодействует с HSPG. Процесс слияния происходит через комбинацию GLUT1 с HSPG/NRP-1, после чего ядро, содержащее вирусную РНК, доставляется в цитоплазму клеток-мишеней. После обратной транскрипции HTLV-1 встраивает свой геном в генома хозяина, чтобы сформировать провирус, окруженный двумя LTR на 5' и 3' концах. Затем провирус транскрибируется и кодирует структурные, регуляторные и вспомогательные белки. Впоследствии вирусные геномные РНК и Gag, Env и Gag-Pol белки переносятся в плазматическую мембрану, чтобы начать собираться в зрелую вирусную частицу.
Вирусы гепатита
Вирус гепатита В (HBV) и вирус гепатита С (HCV) представляют собой несвязанные вирусы, имеющие тропность к гепатоцитам и реплицирующиеся в них. Около 2 миллиардов человек инфицированы HBV, и более 350 миллионов человек становятся хроническими носителями. Только 5-10% взрослых, которые приобретают инфекцию, становятся хроническими носителями, у 30% развиваются прогрессирующие хронические заболевания печени: гепатит, фиброз, цирроз и, наконец, гепатоцеллюлярная карцинома (HCC).
Патогенез. HCC, индуцированная посредством HBV и HCV, развивается в области воспаления и регенерации, что является результатом хронического повреждения печени, а это в свою очередь указывает на то, что патогенез HCC иммунно-опосредован. Эти вирусы постоянно реплицируются в культуре клеток без явных повреждений и смерти клеток, подразумевая, что они являются не цитопатическими. Постоянная репликация вируса является фактором риска для образования HCC, потому что воспаление часто приводит к длительным хроническим заболеваниям печени CLD без выведения вируса. В этом контексте HBV и HCV генерируют белки, подавляющие иммунитет, который должен контролировать хронические инфекции. HBV- и HCV-кодированные белки изменяют экспрессию гена хозяина и клеточный фенотип, что является признаком рака. Эти изменения способствуют независимой от фактора роста пролиферации, сопротивлению к ингибированию роста, инвазии тканей и метастазированию, ангиогенезу, перепрограммированию энергетического метаболизма и устойчивости к апоптозу. Хроническое воспаление также способствует генетической нестабильности в клетках опухоли. Вклад HBV в развитие HCC включает в себя экспрессию гена гепатита Bx (HBx) и, возможно, усеченные пре-S или S-полипептиды карбоксимицина; основной белок и неструктурные (NS) белки NS3 и NS5A HCV способствуют онкогенной трансформации. Изменения в экспрессии гена хозяина, которые способствуют возникновению опухолевого генеза, также, похоже, поддерживают репликацию вируса и/или защиты вирусных гепатоцитов от иммунного опосредованного повреждения и разрушения.

Иммунитет играет важную роль в исходе острых инфекций. Быстрый, сильный и мультиспецифичный ответ против многих белков вирусов гепатита В (HBV) и гепатита C (HCV) приводит к острой инфекции, устраняемой клеткой-хозяином.
Источники
Oncogenes and RNA splicing of human tumor viruses. Masahiko Ajiro & Zhi-Ming Zheng.(September 2014)
Carcinogenic human papillomavirus infection. Mark Schiffman, John Doorbar, Nicolas Wentzensen, Silvia de Sanjosé, Carole Fakhry, Bradley J. Monk, Margaret A. Stanley & Silvia Franceschi.( December 2016)
Human T-cell lymphotropic virus type 1 and its oncogenesis Lan-lan ZHANG, Jing-yun WEI, Long WANG, Shi-le HUANG, Ji-Long CHEN.(2017)
Immunoblastic Lymphoma in Persons with AIDS-Associated Kaposi's Sarcoma: a Role for Kaposi's Sarcoma–Associated Herpesvirus. Eric A Engels M.D., Stefania Pittaluga M.D., Denise Whitby Ph.D., Charles Rabkin M.D., Yoshiyasu Aoki M.D., Elaine S Jaffe M.D. & James J Goedert M.D.(may 2003)
Kaposi’s sarcoma-associated herpesvirus ORF34 is essential for late gene expression and virus production. Mayu Nishimura, Tadashi Watanabe, Syota Yagi, Takahiro Yamanaka & Masahiro Fujimuro( march 2017)
Саркома Капоши: патогенез и основы терапии Н.С. Потекаев, Г.А. Паньшин, Н.П. Теплюк, Н.С. Арсентьев, Е.Ю.,Вертиева,Г.И. Махов, Е.С. Сизова, В.С. Пауков, М.Г. Карташова. (март 2013)
Human T-Cell Lymphotropic Virus: A Model of NF-κB-Associated Tumorigenesis. Zhaoxia Qu and Gutian Xiao(June 2011)
Immune responses and immunopathology in acute and chronic viral hepatitis. Eui-Cheol Shin, Pil Soo Sung & Su-Hyung Park(July 2016)
Pathogenic mechanisms in HBV and HCV-associated hepatocellular carcinoma. Alla Arzumanyan, Helena M. G. P. V. Reis and Mark A. Feitelson(February 2013) Epstein–Barr virus: more than 50 years old and still providing surprises. Lawrence S. Young, Lee Fah Yap & Paul G. Murray( September 2016)
Медицинская микробиология, вирусология и иммунология. В.В. Зверев, А. С. Быков.
Такая зависимость впервые была показана в 1908 г., когда датские ученые Эллерман и Банг нашли, что у цыплят возбудитель лейкоза может быть многократно пропущен через бактериальные фильтры и не теряет при этом своей агрессивности. В 1910 г. американец Пейтон Раус установил связь вирусов с саркомой кур. В 1934 г. его соотечественник Р. Шоуп выделил вирус из опухоли бородавок, или папиллом, кроликов. В 1936 г. Дж. Биттнером был открыт вирус рака молочной железы мышей, и 1951 г. Л. Гроссом — вирус лейкоза мышей, в 1964 г. В. Ярретом — вирус лейкоза домашних кошек, а затем и вирус лейкоза обезьян. В изучение последнего большой вклад внесли советские ученые под руководством академика АМН СССР Б. А. Лапина.
Содержание
Введение…………………………………………………………. 3-4
Особенности онкогенных вирусов…………………………..……4-6
Механизмы действия онковирусов……………………………. 6-9
Заключение………………………………………………………..9-11
Список использованной литературы…………………………. ….12
Вложенные файлы: 1 файл
онковирусы.doc
Неопластическое превращение клеток - сложный, многоступенчатый процесс, ключевым элементом которого является онкоген в составе генома РНК- и ДНК-содержащих опухолеродных вирусов. Он обусловливает последовательную цепь превращений, приводящих к опухолевой трансформации клетки.
Отличительная особенность онкогенов ретровирусов заключается в том, что они не нужны для саморепродукции вируса, их последовательности не родственны последовательностям структурных вирусных генов (gag, pol и env), но зато близкородственны нуклеотидным последовательностям нормальных клеток.
Онкогены ДНК-содержащих опухолеродных вирусов, наоборот, в процессе эволюции стали необходимы для репликации вируса, однако их последовательности негомологичны клеточным последовательностям, т. е, вероятно, они превратились в вирусные и перестали быть клеточными.
Можно заключить, что вирусные онкогены имеют клеточное происхождение и возникли в результате редких трансдукций определенных клеточных последовательностей вирусами.
Молекулярно-биологическими методами установлено, что нормальные клетки позвоночных действительно содержат семейство таких эволюционно консервативных последовательностей (протоонкогенов). Очевидно, они непосредственно участвуют в дифференцировке клеток или других жизненно важных процессах и могут вовлекаться в онкогенез, обусловливая, по-видимому, определенные этапы опухолевой трансформации клеток.
Показано, что вирусные онкогены и клеточные протоонкогены, имеющие общие специфические последовательности, не изогенны. Они отличаются друг от друга по рассеянным точечным мутациям, наличию делеций и уникальных кодирующих участков, которые отсутствуют у их клеточных гомологов, например, протоонкогенов c-src, c-myc, c-ras, c-fps и др.
Естественно, что продукты вирусных и клеточных онкогенов также отличаются друг от друга, и, следовательно, функционально они могут быть различны: вирусные онкобелки трансформируют клетки, а продукты протоонкогенов могут выполнять разные нормальные функции как в эмбриогенезе, так и при физиологических процессах в организме.
Отмечено, что некоторые клеточные онкогены экспрессируются на определенных стадиях онтогенеза и при регенерации органов и тканей. Так, при регенерации печени в 2,5 — 4 раза увеличивается количество транскриптов c-Ha-ras, C-Ki-ras и с-myc-специфических РНК.
Причем высокий уровень экспрессии онкогенов в онтогенезе, например, гена c-src, по данным A. Bernekow и М. Gebler (1984), не всегда связан с пролиферацией, а, возможно, играет определенную роль на ранних стадиях дифференцировки мезенхимальных клеток или же осуществляет контроль метаболизма, в частности, как было показано J. G. Spivack и соавт. (1984), прогестерона, индуцирующего созревание ооцитов. Иными словами, протоонкогены, содержащие онкородственные последовательности с вирусными трансформирующими генами, экспрессируются в нормальных клетках в соответствии с нормальными функциями.
До сих пор не существует функциональных или достоверных доказательств того, что именно только эти клеточные гены вызывают злокачественные новообразования у животных, не зараженных вирусами, которые содержат онкогены. Поэтому остается неясным, имеют ли протоонкогены трансформирующий потенциал или являются только прародителями вирусных онкогенов.
Не решен также вопрос о соотношениях вирусных и клеточных онкогенов, сохраняющих транспозонную природу, в процессе неопластической трансформации клеток. Нельзя исключить, что существование участков клеточных ДНК с повышенной склонностью к транслокации, в которых расположены некоторые клеточные онкогены, имеют важное физиологическое значение для экологической и, в частности, иммунологической адаптации организмов.
Anders Valhne
Легче в биологии сделать открытие, чем получить за это признание.
Peyton Rous
В ХХ веке существовали три теории развития рака. В возникновении злокачественных опухолей активно обсуждалась роль канцерогенов, онкогенных мутаций и онкогенных вирусов. Вирусную теорию развития рака не признавали более 40 лет.
Фильтрующиеся вирусы и рак
Фильтрующийся вирус был открыт профессором Д. И. Ивановским в 1892 г. Работая в Никитском ботаническом саду, он установил причину развития мозаичной болезни листьев табака. Используя фильтр Шамберлана-Пастера, содержащий неглазурованный фарфор, ученый получил бесклеточный фильтрат из больных табачных листьев и заразил этим фильтратом здоровые растения. Поскольку фильтрат не содержал клеток, профессор сделал вывод, что мозаичная болезнь вызывается токсинами или ядами. Позже выяснилось, что причиной болезни были вирусы, получившие название фильтрующихся. Формально Д. И. Ивановский был прав, поскольку латинское слово virus переводится на английский как poison – яд.
В 1908 г. датским ученым Vilhelm Ellerman и Oluf Bang впервые удалось вызвать лейкоз у птиц, заразив их вирусом эритробластоза. Этими исследованиями был заложен фундамент для работ Пейтона Роуса.
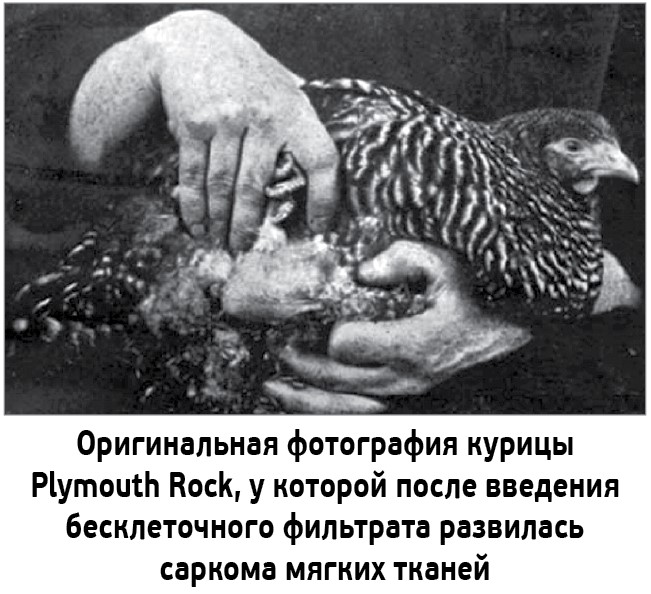
В 1909-1911 гг. Роус работал в Университете Рокфеллера, США. Случайно ему стало известно, что на одном из фермерских хозяйств все куры породы Plymouth Rock болеют саркомой мягких тканей. Такая же опухоль быстро появлялась и у здоровых птиц, попавших на ферму. Повторив эксперимент с фильтром Шамберлана-Пастера, Роус получил из опухоли больной курицы бесклеточный фильтрат, с помощью которого смог вызвать развитие саркомы de novo у здорового цыпленка, не перевивая ему культуру злокачественных клеток. Ученый объявил, что причиной развития опухоли у исследуемых птиц были вирусы (Rous P. A transmissible avian neoplasm. (Sarcoma of the common fowl). J. Exp. Med. 1910; 12: 696-705).
Год, когда Роус начал свои эксперименты, можно считать началом новой науки – вирусологии опухолей человека. Однако медицинский мир посчитал эксперименты Роуса курьезными и не научными. Сторонники теории вирусного канцерогенеза длительное время подвергались критике, в том числе и потому, что на протяжении десятилетий никак не удавалось найти вирус, вызывающий опухоли не у животных, а у человека. Роус впервые номинировался на Нобелевскую премию еще в 1926 г., но получил всемирное признание только в 1966-м, за 4 года до смерти.
Онкогенные вирусы человеческого рака

Поиски вирусов человеческого рака были неудачными, поскольку, как теперь известно, онкогенные вирусы в клетках опухоли не реплицируются и не образуют вирионы, их невозможно обнаружить обычными вирусологическими методами. Первый вирус опухоли человека – вирус Эпштейна-Барр (EBV), также известный как вирус герпеса человека 4 типа (HHV4), – был обнаружен только через 53 года после экспериментов Роуса. Энтони Эпштейн, Берт Ачонг и Ивонна Барр в 1964 г. использовали электронную микроскопию для обнаружения частиц этого вируса в клеточных линиях лимфомы Беркитта.
Обнаружение EBV было случайным, так как большинство опухолевых клеток лимфомы содержат вирусную ДНК в непередаваемой эписомальной форме. С момента этих исследований прошло еще 30 лет, прежде чем Международным агентством по исследованию рака было официально объявлено, что EBV является доказанным человеческим канцерогеном.
Сегодня список вирусов, вызывающих опухоли у человека, остается небольшим. Известны семь вирусов человеческих опухолей: EBV, вирус гепатита B (HBV), вирус гепатита C (HCV), T‑лимфотропный вирус человека 1 типа (HTLV‑1), семейство папилломавирусов человека (HPV), вирус герпеса, ассоциированный с саркомой Капоши (KSHV), и полиомавирус клеток Меркеля (MCPyV). Они были открыты на протяжении 1964-2008 гг.
Онкогенные вирусы имеют различный клеточный тропизм. EBV является причиной большинства лимфом Беркитта, рака носоглотки, лимфопролиферативных заболеваний, некоторой части болезни Ходжкина, неходжкинских лимфом и лимфом желудочно-кишечного тракта. HBV ассоциирован с развитием гепатоцеллюлярной карциномы, а HCV является флавовирусом и отвечает за развитие рака печени и некоторых лимфом. HTLV-I вызывает Т‑клеточный лейкоз у взрослых, HPV – аногенитальные раки, рак шейки матки, полового члена, раки головы и шеи. KSHV является причиной саркомы Капоши, некоторых видов лимфом и редкого доброкачественного заболевания лимфатических узлов, проявляющегося гигантской ангиофолликулярной гиперплазией (болезнь Кастлемана). Последний член группы вирусов человеческого рака, внутриклеточный полиомавирус Меркеля (MCV), длительное время персистирует в фибробластах кожи и вызывает одноименную болезнь – нейроэндокринную карциному Меркеля.
Постоянно предпринимаются попытки расширить этот список, однако роль новых онкогенных вирусов в канцерогенезе человеческого рака остается неясной и противоречивой.
Молекулярные особенности онкогенных вирусов
Онкогенные вирусы не попадают ни в один из семи классов вирусов по классификации Балтимора (1971). Эта классификация разделяет вирусы на группы в зависимости от типа нуклеиновой кислоты и способов ее репликации (ДНК, РНК, одноцепочечная, двуцепочечная). Отсутствует четкая молекулярная характеристика, которая указывала бы на принадлежность к вирусам человеческого рака.
Так, EBV и KSHV являются крупными двуцепочечными ДНК‑вирусами. Семейство HPV представлено небольшими онкогенными ДНК‑вирусами с меньшими геномами и малым количеством многофункциональных онкопротеинов. MCPyV – небольшой двуцепочечный ДНК‑вирус. HBV является маленьким ДНК‑вирусом с ретровирусными функциями, HCV – одноцепочечным вирусом с положительной цепью РНК. HTLV‑1 является сложным экзогенным ретровирусом человека.
Несмотря на сложные молекулярные различия, все онкогенные вирусы имеют два общих признака: они способны вызывать хроническую бессимптомную инфекцию и сосуществуют с хозяином в течение длительного периода времени.
Молекулярное паразитирование вирусов
Внутриклеточные мишени вирусов человеческого рака хорошо изучены. Это пролиферативные сигнальные пути опухолевых супрессоров RB1 и p53, теломеразная обратная транскриптаза, путь PI3K/AKT/mTOR, транскрипционный ядерный фактор κB, β-катенин (также известный как CTNNB1) и пути передачи сигналов интерферона. Данные молекулярные события являются не только важными механизмами регуляции клеточной пролиферации, но и сигнальными путями врожденного иммунного ответа, что определяет их роль в реализации вирусного канцерогенеза.
Основным механизмом защиты клетки от вируса является апоптоз. В ответ на вирусную инфекцию клетки активируют гены-супрессоры RB1 и p53 и останавливают клеточный цикл в контрольных точках (check-point). В дальнейшем могут включиться сигнальные пути гибели клеток.
Однако целью вирусов является не гибель клеток, а репликация собственной ДНК и эффективное воспроизводство. Это становится возможным только в том случае, если клетка завершит клеточный цикл. С целью предупреждения преждевременной гибели клеток опухолевые вирусы разработали уникальные механизмы инактивации белков-супрессоров RB1 и p53, блокада которых препятствует уходу клетки в апоптоз, способствует развитию иммунной толерантности и дальнейшему продвижению клетки к митотической фазе клеточного цикла, где произойдет репликация вирусной ДНК. Таким образом, функция вирусных онкогенов заключается в блокировании генов-супрессоров, продвижении клетки в клеточном цикле и стимулировании продуктивной репликации вирусов.
Обычно инактивация вирусом белков-супрессоров RB1 и p53 не приводит к развитию рака. При хронической вирусной персистенции опухоль может возникнуть только при сопутствующей иммуносупрессии и появлении дополнительных онкогенных мутаций в клетках хозяина.
Вирусные канцерогены
Принято считать, что вирусы могут инициировать развитие рака двумя путями – с помощью прямых и косвенных канцерогенов. Прямые вирусные канцерогены присутствуют в каждой раковой клетке и экспрессируют по меньшей мере один транскрипт для поддержания фенотипа опухолевых клеток. Так происходит при раках, ассоциированных с вирусами HPV, MCV, EBV и KSHV.
Косвенные канцерогены вызывают рак посредством хронической инфекции и воспаления, которые протекают в тканях латентно и не проявляются клинически. Клинические симптомы отсутствуют даже в случаях активной репликации цитолитического вируса, как это происходит во время продромальной фазы ВИЧ- или HCV‑инфекции.
Вирусный онкогенез
Равновесие, достигаемое вирусами и соматическими клетками, может оказаться нестабильным. На фоне индуцированной вирусом иммуносупрессии и при внезапном появлении новых онкогенных мутаций происходит инициация начальных стадий канцерогенеза. Вирусные онкогены могут вызвать анэуплоидность и нестабильность генома клетки, что способствует ее злокачественной трансформации.
Геномная нестабильность, например, часто наблюдается при неоплазиях шейки матки, связанных с HPV‑вирусами высокого онкогенного риска. Онкопротеины HPV Е6 и Е7 дестабилизируют клеточный геном, вызывают повреждение и нарушают репарацию ДНК, что увеличивает вероятность приобретения дополнительных онкогенных мутаций.
Следует отметить, что роль вирусов важна на самых ранних стадиях злокачественной трансформации клетки. В опухолевой ткани вирусы долго не сохраняются и поэтому на поздние стадии канцерогенеза, в том числе на стадии метастатической прогрессии, влияния не оказывают.
Все вирусы человеческого рака обладают способностью образовывать вирионы в любые моменты своего естественного жизненного цикла, однако в тканях опухолей репликация продуктивного вируса (литическая репликация) отсутствует. Эта особенность является общей для всех вирусов человеческого рака.
По мере созревания опухоль теряет вирусные гены и белки. При латентной вирусной персистенции в опухолевой ткани не происходит размножение вирусов и не образуются инфекционные вирусные частицы. В клетках рака вирус сохраняется в виде голой нуклеиновой кислоты, часто в виде плазмид или эписом.
Иммунная система и вирусы человеческого рака
За последнее десятилетие накопилось большое количество свидетельств о важной роли врожденного иммунитета в вирусном онкогенезе. Онкогенные вирусы способны уклоняться от механизмов иммунной системы хозяина. Для этого они применяют сложные стратегии выживания. Вирусные латентность и персистенция проявляются отсутствием клинических признаков хронической инфекции, потерей вирусных генов и белков, хранением информации в виде нуклеиновых кислот или эписом.
Уклонение от контролирующего влияния иммунной системы хозяина, с помощью которого вирусные онкогены обеспечивают вирусную латентность и одновременно способствуют выживанию инфицированной клетки, является обязательным условием развития опухолей вирусной этиологии.
Многие ключевые эффекторные белки являются общими как для супрессоров опухолей, так и для сигнальных сетей врожденного иммунного надзора. Оба процесса инициируют остановку клеточного цикла и активируют первичные апоптотические пути. Блокада супрессоров вирусными онкогенами способствует уклонению от иммунитета и непреднамеренно подвергает инфицированную клетку риску раковой трансформации.
Генерализованная иммуносупрессия (медикаментозная или во время процесса старения) сопровождается потерей иммунного надзора за специфическими вирусными Т‑клеточными эпитопами и всегда связана с риском развития рака.
Роль иммунной системы в развитии злокачественной опухоли хорошо видна на примере эволюции саркомы Капоши в человеческом обществе. Саркома Капоши впервые была описана в 1872 г. В XIX и первой половине XX века частота заболевания составляла менее 3 случаев на 1 млн населения в год. После эпидемии СПИДа она возросла в десятки тысяч раз. Росту частоты саркомы Капоши в XX столетии также способствовало развитие органной трансплантации и методов лекарственной профилактики острого отторжения, ведущих к стойкой депрессии иммунной системы у реципиентов донорских органов. Люди с пересаженными органами относятся к группе высокого онкологического риска, но в первую очередь – риска развития саркомы Капоши, имеющей вирусную этиологию.
Злокачественная клетка – тупиковое событие в эволюции вирусов человеческого рака
Люди, как и большинство сложных многоклеточных, являются химерами для многочисленных вирусов. Онкогенные вирусы эволюционировали, чтобы длительно сохраняться в клетках хозяина. В свою очередь, у человека развились механизмы для поддержания целостности и стабильности инфицированных вирусом клеток, которые должны выполнять жизненно важные функции в организме.
На протяжении миллионов лет сложились синергические отношения между некоторыми онкогенными вирусами и клетками человека. Эти отношения предполагают установление равновесия, в результате которого персистенция вируса способствует репликации вирусной ДНК, однако не приводит к гибели клетки, поскольку вирусы блокируют внутриклеточные пути апоптоза, что способствует завершению клеточного цикла. Врожденная иммунная система человека, предназначенная для защиты генома хозяина от встраивания чужеродных вирусных последовательностей ДНК, также инактивируется вирусами через гены-супрессоры, что способствует латентной вирусной персистенции.
Судьба хозяина и патогена зависит от равновесия, при котором с обеих сторон достигается контроль сигнальных путей роста, механизмов поддержания стабильности генома и иммунного надзора. Обычно молекулярный паразитизм вируса не приводит к развитию рака. Лишь у очень небольшой части людей, инфицированных онкогенными вирусами, возникает злокачественная опухоль. Эти больные никогда (или почти никогда) не служат источниками передачи и распространения вирусов в человеческой среде. Однако в случае кумулятивных или случайных событий, вызывающих повреждение ДНК или подавление иммунной системы, хрупкий баланс между вирусом и клеткой может нарушиться. Следствием этого является неконтролируемая клеточная пролиферация.
При злокачественной трансформации отношения между вирусом и клеткой меняются. В инфицированных раковых клетках не улучшается способность вирусов к репликации и не увеличивается их способность к размножению, как это происходит при воздействии на клетки цитолитических вирусов, вызывающих острую инфекцию у человека. По мере созревания опухолевые клетки вообще утрачивают вирусную ДНК, из-за чего обнаружить следы вируса в злокачественных клетках опухоли обычными вирусологическими методами не удается. С эволюционной точки зрения рак для вирусов является тупиковым событием, биологическим несчастным случаем. При злокачественной трансформации клеток репликация вирусной ДНК в них прекращается и вирусы утрачивают способность сохранять и передавать свою генетическую информацию.
Таким образом, большинство классов вирусов не запрограммированы эволюцией вызывать рак у человека. Однако опухолевые вирусы все же кодируют человеческие онкогены, активируют сигнальные пути и способствуют избыточной пролиферации инициированных клеток. Пролиферация инфицированной клетки является не целью, а побочным продуктом влияния вируса.
Выводы
На протяжении большей части XX века вирусная теория рака, основы которой в 1909 г. заложил П. Роус, подвергалась острой критике. Обычные вирусологические методы не обнаруживали следов вирусов в клетках человеческих опухолей, и поэтому роль вирусов в канцерогенезе отвергалась. В науке господствовала догма Роберта Коха, согласно которой инфекционная причина болезни признавалась только в том случае, если возбудителя можно было обнаружить в клетках или тканях больного. По этой же причине многие десятилетия были потрачены впустую на поиск бактерий, вызывающих рак.
После исследований Renato Dulbecco, Howard Temin и David Baltimore (Нобелевская премия 1975 г.) противоречия между канцерогенной, вирусологической и генетической теориями возникновения опухолей были наконец устранены. Открытие феномена обратной транскрипции, обнаружение вирусных онкогенов и генов-супрессоров привели к созданию теории соматических мутаций, которая является фундаментальной в современной онкологии.
Взаимоотношения между соматическими клетками эукариот и онкогенными вирусами установились еще в эпоху метазоа и подчиняются законам эволюции. Вирусы демонстрируют различный клеточный тропизм и молекулярный паразитизм, который проявляется блокадой белков-супрессоров, ингибирующих апоптоз, уклонением от контроля иммунной системы, нелитической репликацией вирусной ДНК и длительной латентной персистентностью.
Принято считать, что около 20% всех человеческих опухолей имеют вирусную этиологию. Вирусология опухолей человека имеет 110-летнюю историю, но только в последнее время эта наука приобрела практическое значение. В первую очередь это связано с программами первичной профилактики рака. Вакцины против онкогенных штаммов HPV привели к резкому снижению заболеваемости раком шейки матки во многих странах мира, где эти программы носят государственный характер. Вакцинация от HBV привела к снижению частоты первичного рака печени. Особенно это стало заметно в последние годы в странах с высокой заболеваемостью этим видом рака. Эффективной профилактикой первичной гепатоцеллюлярной карциномы являются современные стратегии противовирусной терапии, направленные против HCV. К сожалению, несмотря на то что EBV и KSHV являются доказанными биологическими канцерогенами и вызывают большое количество злокачественных лимфом, вакцин к ним до сих пор нет.
Развитие омических технологий (в частности, методов секвенирования нового поколения) открыло новые перспективы для вирусологии опухолей человека. Сегодня появились большие возможности для обнаружения новых вирусов человеческого рака. Например, глубокое секвенирование и профилирование экспрессии генов недавно привело к открытию MCPyV – хронологически последнего в семействе вирусов человеческого рака. Можно предсказать, что в ближайшее время будут обнаружены и другие вирусы, которые отвечают за развитие злокачественных опухолей у человека.
Проводятся исследования и ожидается создание лекарственных препаратов, блокирующих пути вирусного канцерогенеза. Такими могут быть препараты для иммунной терапии, направленной против антигенов онкогенных вирусов.
Современная вирусология опухолей человека, использующая методы молекулярной биологии, позволит на новом уровне изучить механизмы вирусного канцерогенеза, создать программы массовой профилактики, диагностики и лечения вирусного рака.
СТАТТІ ЗА ТЕМОЮ Інфекційні захворювання
Інфекція нижніх дихальних шляхів (ІНДШ) – гостре захворювання (триває протягом ≤21 дня) з основним симптомом – кашлем і щонайменше одним з інших симптомів з боку дихальних шляхів (відходження мокротиння, задишка, хрипи чи дискомфорт / біль у грудях), що не має альтернативних пояснень (наприклад, астми/синуситу). ІНДШ включає різні захворювання, як-от гострий бронхіт (ГБ), грип, позалікарняна пневмонія (ПЛП), загострення хронічного обструктивного захворювання легень (ХОЗЛ) і бронхоектатичної хвороби (БЕХ).
Читайте также: